Nutrient Digestibility and Metabolizable Energy Content of Mucuna pruriens Whole Pods Fed to Growing Pelibuey Lambs
Article information
Abstract
The nutrient digestibility, nitrogen balance and in vivo metabolizable energy supply of Mucuna pruriens whole pods fed to growing Pelibuey lambs was investigated. Eight Pelibuey sheep housed in metabolic crates were fed increasing levels of Mucuna pruriens pods: 0 (control), 100 (Mucuna100), 200 (Mucuna200) and 300 (Mucuna300) g/kg dry matter. A quadratic (p<0.002) effect was observed for dry matter (DM), neutral detergent fibre (aNDF), nitrogen (N) and gross energy (GE) intakes with higher intakes in the Mucuna100 and Mucuna200 treatments. Increasing M. pruriens in the diets had no effect (p>0.05) on DM and GE apparent digestibility (p<0.05). A linear reduction in N digestibility and N retention was observed with increasing mucuna pod level. This effect was accompanied by a quadratic effect (p<0.05) on fecal-N and N-balance which were higher in the Mucuna100 and Mucuna200 treatments. Urine-N excretion, GE retention and dietary estimated nutrient supply (metabolizable protein and metabolizable energy) were not affected (p>0.05). DM, N and GE apparent digestibility coefficient of M. pruriens whole pods obtained through multiple regression equations were 0.692, 0.457, 0.654 respectively. In vivo DE and ME content of mucuna whole pod were estimated in 11.0 and 9.7 MJ/kg DM. It was concluded that whole pods from M. pruriens did not affect nutrient utilization when included in an mixed diet up to 200 g/kg DM. This is the first in vivo estimation of mucuna whole pod ME value for ruminants.
INTRODUCTION
Currently, there is a renewed interest to evaluate feed potential of M. pruriens in ruminant animals (Castillo-Caamal et al., 2003; Matenga et al., 2003; Chikagwa-Malunga et al., 2009a, b, c, d; Loyra-Tzab et al., 2011, García-Galvan et al., 2012).
Mucuna whole pods or beans alone could be used as supplement to ruminants fed poor quality roughage diets in many tropical countries. This option would be particularly important during the dry season when there is a reduction in productive capacity of animals due to inadequate availability and low quality of pastures (Castillo-Caamal et al., 2003; Matenga et al., 2003).
Although anti-nutritional factors have been detected in M. pruriens (particularly in beans), there is no evidence of a detrimental effect of these compounds when M. pruriens is fed to ruminants in large quantities (Castillo-Caamal et al., 2003; Mendoza-Castillo et al., 2003; Pérez-Hernández et al., 2003).
The M. pruriens bean contains a high amount of crude protein (CP) while its husk has a lower CP and a high content of lignin and fiber (Ayala-Burgos et al., 2003; Sandoval-Castro et al., 2003). The M. pruriens pods contain 56% grain and 44% husk in terms of dry matter (DM) (Sarmiento et al., 2006 unpublished data). Ayala-Burgos et al. (2003) reported effective DM degradability in sacco being 81% and 53% for bean and husk respectively. Higher values were obtained by Sandoval-Castro et al. (2003) by an in vitro digestibility technique (97.94 and 78.96% for bean and husk respectively). The organic matter (OM) digestibility from bean and husk were 96.02 and 78.85% respectively. Although, degradability values from the husk are inferior, they are still higher than those of common grasses used for ruminant livestock in many tropical areas of Mexico (Ayala-Burgos et al., 2003; Sandoval Castro et al., 2003).
Studies evaluating nutrient digestibility or nitrogen retention from pods or bean alone of M. pruriens in ruminants are scarce (Chay-Canul et al., 2009; Chikagwa-Malunga et al., 2009d; Loyra-Tzab et al., 2011). Information available on metabolizable energy (ME) concentration from Mucuna bean and husk has been estimated from results obtained with in vitro digestibility techniques. Thus, for the beans Burgos et al. (2002) and Sandoval-Castro et al. (2003) reported an ME value for ruminants of 11.9 and 13.9 MJ/kg DM respectively and for husk 11.4 MJ/kg DM (Sandoval-Castro et al., 2003). These results suggest that M. pruriens could be employed as a source of nitrogen (N) and energy by the rumen microorganisms as there are no effects upon rumen microbial activity (Chikagwa-Malunga et al., 2009a).
It would be attractive for farmers to use the whole pod rather than only the bean, while discarding the husk, since the husk appear to have high digestibility. It is likely that including mucuna whole pod in ruminant diets will not cause major changes in their performance as compared with traditional feeds (García-Galván et al., 2012). However, in order to incorporate mucuna in commercial formulations, an estimate of energy retention and metabolizable energy values are needed. In a previous work (Loyra-Tzab et al., 2011) in vivo estimation of mucuna bean ME content was reported. However, a ME value for mucuna whole pod has not been estimated.
Thus, the objectives of this study were i) to determine apparent digestibility of Mucuna whole pod (DM, N, and gross energy (GE)), ii) to estimate N and GE retention coefficients when the M. pruriens whole pods was incorporated in a diet and iii) to estimate in vivo Mucuna whole pod ME value in diets for growing Pelibuey lambs.
MATERIALS AND METHODS
The experiment was conducted at the Faculty of Veterinary Medicine and Animal Science (University of Yucatan), located in the central region of the state of Yucatan, Mexico (21° 15’ N lat., 83° 32’ long.). The region has an annual average rainfall of 953 mm. Annual average temperature and humidity of 26.5°C and 72% respectively.
The methodology employed was similar to that reported by Loyra-Tzab et al. (2011) where mucuna bean was evaluated. In this experiment mucuna whole pods (husk and bean) were evaluated. The methodology is described below.
Animals
Eight Pelibuey male lambs (29.37±3.24 kg live weight) were housed in metabolic cages allowing a complete separation and recovery of faeces and urine. Lambs were treated for internal parasites by giving an oral dose of albendazole (7.5 mg/kg live weight (LW)) before the start of the trial.
Experimental diets
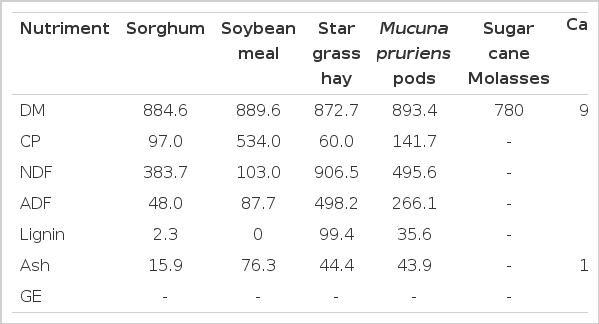
At maturity M. pruriens pods were harvested under tropical conditions of Yucatán, Mexico. Star grass (C. nlenfuensis) was harvested at 4 wk of re-growth. Both star grass and M. pruriens pods were sun dried and ground through a 3 mm sieve. Sorghum grain, soybean meal and CaCO3 were obtained from a commercial company of the region. M. pruriens pod was mixed with the corresponding amounts of a control diet in a horizontal mixer. Control diet consisted of 545.4, 110.1, 312.6, 29.8 and 2.1 g/kg DM of sorghum grain, soybean meal, star grass hay, molasses sugarcane and CaCO3 respectively. Experimental diets consisted of ground M. pruriens pods in the following proportions 0 (control diet); 100, 200 and 300 g/kg DM. Levels were selected to allow the estimation of energy value of Mucuna beans using the methodology of Schneider and Flatt (1975) and taking into account feasible feed formulation inclusion levels. Diets had similar CP and GE contents. The chemical composition of experimental diets is shown in Table 1. Lambs were fed daily ad libitum (at 09:00 h), allowing for a 20% increase over the DM consumed the previous day. Water was freely available.
Feed intake and apparent digestibility
The amounts of both consumed and refused feed were recorded daily and samples were taken for chemical analyses. DM, N and GE intakes were measured as the difference between the amounts offered and those refused. Faeces were collected daily, weighed and samples were kept. All samples of feeds and faeces were stored at −20°C until analysis.
N and GE retention and N balance
Daily urinary excretion was weighed and a proportional sample was taken from each animal. The urine samples were kept at a pH below 3.0, by adding 80 ml of a diluted solution of Sulphuric acid (4 M) per liter. N content of feed, feces and urine were used to estimate N balance and retention. In addition, energy excreted through urine was estimated according to McDonald et al. (2002) from urine-N content as 23 kJ/g N excreted. A constant level of 0.08 of GE was considered to be lost as methane.
Chemical analyses
Daily feed samples and orts were proportionately pooled to obtain a single sample for each period. Samples were analyzed for DM (#7.007), CP (N*6.25) (#2.057) and ash (#7.009) (AOAC, 1980). GE was determined in an adiabatic calorimetric chamber (Parr, 1988). Neutral detergent fiber (aNDF) (Mertens, 2002), acid detergent fiber (ADF) (Van Soest et al., 1991) and lignin (sa) (Robertson and Van Soest, 1981) were also determined. aNDF was analyzed without sulphite, with amylase and ash inclusive. Faecal samples were analyzed for DM, GE and N. In addition, urine-N was determined. The methods utilized were the same as above.
Statistical analysis
A balanced block design (Mead, 1988) with two periods of fifteen days each, ten days for adaptation and five days for measurements were used. On each period, two animals were randomly allocated to each of the four treatments. A total of four animals (replicates) per treatment was obtained after the two experimental periods. One week interval was allowed between both periods where animals were fed the control diets.
Data were analyzed using a generalized linear model (Minitab, 2007). The model considered the effects of treatments (4 Mucuna inclusion levels) and experimental periods were considered as blocking factors (2 periods). Total number of replicates per treatment was 4. Degrees of freedom for experimental error = 11. Linear, quadratic and cubic effects were determined. Significant effects were accepted if p≤0.05. In addition, DM, CP and GE apparent digestibility coefficients, as well as digestible protein (DP), digestible energy (DE) and metabolizable energy (ME) content values from M. pruriens beans were calculated using multiple regression equations (Schneider and Flatt, 1975).
RESULTS
Nutrient intake and digestibility of experimental diets
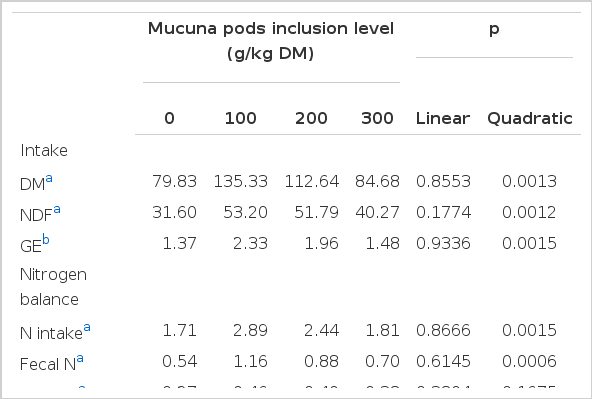
A quadratic (p<0.002) effect was observed for dry matter (DM), neutral detergent fibre (aNDF), nitrogen (N) and gross energy (GE) intakes with higher intakes in the Mucuna100 and Mucuna200 treatments. Increasing M. pruriens in the diets had no effect (p>0.05) on DM and GE apparent digestibility (p<0.05). A linear reduction (p<0.04) in N digestibility was observed as mucuna whole pods increased in the diet. However, no effects were found on estimated nutrient supply (DP, DE and ME) (Table 2).
Nutrient intake, retention and digestibility in diets containing M. pruriens pods
Nutrient retention and nitrogen balance
Although N balance was positive in all treatments, a linear reduction in N digestibility and N retention was observed with increasing mucuna whole pod level. However, urine-N excretion was not affected (p<0.5). These resulted in a quadratic effect (p<0.05) on fecal-N and N-balance which were higher in the Mucuna100 and Mucuna200 treatments,
Nutrient digestibility and energy content of raw M. pruriens pods
Average DM and GE digestibility coefficients of M. pruriens pods were higher than that of control diet being 0.69±0.060 and 0.65±0.058 vs 0.51±0.042 and 0.51±0.040 respectively. However CP digestibility coefficient of M. pruriens pods was lower than that from the control diet (0.46±0.070 vs 0.54±0.046). The estimated DE and ME values for mucuna pods were 11.0±1.08 and 9.7±1.08 MJ/kg DM respectively.
DISCUSSION
Nutrients intake
Inclusion of M. pruriens pods in diets did not have a negative effect in total DM intake possibly due to the similar chemical composition and nutrient digestibility among M. pruriens pods and control diets. Castillo-Caamal et al. (2003) supplemented a basal diet of Pennisetum purpureum using three levels (165, 331 and 497 g/kg LW) of M. pruriens pods and reported a linear increase in total DM intake, without an effect on forage DM intake. On the other hand, Perez-Hernandez et al. (2003) reported a tendency to decrease the digestibility in P. purpureum basal diet when a concentrate was substituted by M. pruriens pods on 50% and 100% level, which was explained to be due to the increase in fiber (NDF) from the pod husk. However, although the diets were formulated to resemble a commercial lamb fattening concentrate diet, the resulting higher NDF concentration (particularly in Mucuna300 diet) appeared to be a limiting factor for DM intake. The reduction in DM intake was not attributed to an anti-nutritional factor effect as the intake level was similar to that achieve with the control diet and no symptoms of toxicity were observed.
Nutrient digestibility
A similar DM and GE digestibility between all experimental diets was expected due to their similar nutrient composition (Table 2). Although Ayala-Burgos et al. (2003) indicated that according to its chemical composition M. pruriens husk was classified as a roughage. The in vitro and in sacco DM degradability of Mucuna husk (0.79 and 0.53 respectively; Ayala-Burgos et al., 2003; Sandoval-Castro et al., 2003) is higher than those from tropical grasses (Mero and Udén, 1997; Mero and Udén, 1998a). This was in agreement with previous findings on legume pod digestibility (Pieltain et al., 1996; Bruno-Soares and Abreu, 2003). Hence, DM digestibility seems not to be affected by including the husk fraction of legume pods making feasible its routine use for feeding ruminants.
Digestibility, retention and supply of nutrient from diets containing M. pruriens pods
DM and GE digestibility of diets formulated with M. pruriens pods were higher than those from most diets based on tropical grasses. Although its N digestibility was lower than those from diets based on young (re-growth) grass, it was similar or higher to values obtained with mature tropical grasses (Mero and Udén, 1998a; Archimède et al., 2000; Mendoza and Sandoval, 2003).
Digestibility and N retention of Mucuna bean diets has been previously compared with diet containing soybean meal with satisfactory results (Chikagwa-Malunga et al., 2009d; Loyra-Tzab et al., 2011; García-Galvan et al., 2012).
Although a positive N balance was observed among all treatment, similar to previous results (Chikagwa-Malunga et al., 2009d; Loyra-Tzab et al., 2011), N balance was lower at higher inclusion levels. It is possible that soybean meal provides a better amino acid balance as suggested by Chikagwa-Malunga et al. (2009d) but it is also possible the result of accounting L-dopa as a protein-N source. As L-dopa would not accumulate in blood or tissue (Chikagwa-Malunga et al., 2009c), nor utilized for protein accretion, it would be excreted and result in lower N digestibility, MP supply and retention (Table 2). Although as with any other protein rich feed, efficiency of N utilization might also be influenced by diet presentation (e.g. particle size) (Chay-Canul et al., 2009), and grinding whole pods is likely to result in a larger particle size than grinding beans alone.
In vivo metabolizable energy value
To our knowledge, this is the first report of an in vivo ME value for Mucuna whole pod. Its 9.7 MJ/kg DM value compares favorably with many green crops such as rape, maize, sugarcane and lucerne, as well as with other feed such as clover hay (McDonald et al., 2002). It is lower than an estimation based on ME values for bean and husk when these fraction where evaluated as separated fractions (Sandoval-Castro et al., 2003), possibly due to a lower digestibility when incorporated in whole diets. Additionally, in vitro and in situ digestibility estimates represent the potential a feed can achieve which usually are higher than in vivo measurements as samples cannot escape rumen digestion. Nevertheless current estimate provides a value which can be readily incorporated into commercial formulation as has been validated by García-Galván et al. (2012), who found this ME value reliable for feed formulation in growing lambs.
CONCLUSION
Nutrient apparent digestibility and retention in diets incorporating M. pruriens whole pods are similar to those from diets made from conventional feedstuff when included in diets for lambs up to 200 g/kg DM without detrimental effects on nutrient digestibility and retention by growing lambs. The ME value of M. pruriens was estimated to be 9.7 MJ/kg DM.