Evaluation of Anti-SE Bacteriophage as Feed Additives to Prevent Salmonella enteritidis (SE) in Broiler
Article information
Abstract
This experiment was conducted to evaluate anti-Salmonella enteritidis (anti-SE) bacteriophage as feed additives to prevent Salmonella enteritidis in broilers. The experimental diets were formulated for 2 phases feeding trial, and 3 different levels (0.05, 0.1 and 0.2%) of anti-SE bacteriophage were supplemented in basal diet. The basal diet was regarded as the control treatment. A total of 320 1-d-old male broilers (Ross 308) were allotted by randomized complete block (RCB) design in 8 replicates with 10 chicks per pen. All birds were raised on rice hull bedding in ambient controlled environment and free access to feed and water. There were no significant differences in body weight gain, feed intake and feed conversion ratio (FCR) at terminal period among treatments (p>0.05). Relative weights of liver, spleen, abdominal fat and tissue muscle of breast obtained from each anti-SE bacteriophage treatment were similar to control, with a slightly higher value in anti-SE bacteriophage 0.2%. In addition, a numerical difference of glutamic-oxaloacetic transaminase (GOT), glutamic-pyruvic transaminase (GPT) and LDL cholesterol level was observed in the 0.2% anti-SE bacteriophage application even though blood profiles were not significantly affected by supplemented levels of anti-SE bacteriophage (p>0.05). In the result of a 14 d record after Salmonella enteritidis challenge of 160 birds from 4 previous treatments, mortality was linearly decreased with increasing anti-SE bacteriophage level (p<0.05), and Salmonella enteritidis concentration in the cecum was decreased with increasing levels of anti-SE bacteriophage (p<0.05). Based on the results of this study, it is considered that supplementation of 0.2% anti-SE bacteriophage may not cause any negative effect on growth, meat production, and it reduces mortality after Salmonella enteritidis challenge. These results imply to a possible use of anti-SE bacteriophage as an alternative feed additive instead of antibiotics in broilers diet.
INTRODUCTION
In general, Salmonellosis is a severe disease in humans caused by Salmonella bacteria in contaminated food. Salmonella enteritidis is a genetically homogeneous serotype of Salmonella, and it is well known as one of the most common types in food poisoning (Hudson et al., 2001; Chung et al., 2003; Ging et al., 2007). Chickens and eggs are frequently reported as the cause of outbreaks of Salmonellosis (ST Lous et al., 1988; Molbak and Neiman, 2002), and outbreaks of Salmonella bacteria are often detected in broiler flocks. In addition, once broilers are exposed to Salmonella enteritidis, the bacteria quickly spread to the entire poultry flock (Altekruse et al., 2006; Foley et al., 2008; Berrang et al., 2009). The feces from broilers infected with Salmonella enteritidis contaminated an entire chicken slaughterhouse (Shah et al., 2011). From these aspects, prevention of Salmonella enteritidis infection is an important issue in maintaining food safety particularly in the poultry industry. In an attempt to reduce disease including Salmonella enteritidis in poultry, antibiotics were provided as a tool for effective prevention. However, in the recent years, poultry producers are becoming interested in alternative materials rather than antibiotics in animal feed because of public awareness of bacterial antibiotic resistance arising in products from animal origin (Huang et al., 2007; Li et al., 2008).
A bacteriophage or phage is a kind of virus that infects only bacteria (Calton, 1999). They can be observed widely in nature and can be isolated from sewage. Bacteriophage can attach to bacterial cells with their tail fibers and inject the required amount of components for bacteriophage replication into the bacterium. Several studies indicated that Salmonella can be controlled by the use of bacteriophage (Barrow et al., 1987; Berchieri et al., 1991; Sklar and joerger, 2001). In Korea the use of antibiotics in animal feed was prohibited from July 1, 2011, consequently many f poultry producers are interested in bacteriophage as an alternative feed additive to dietary antibiotics for controlling Salmonella infection. A wide variety of methods for bacteriophage application such as air sac inoculation, drinking supply, aerosol spray, and their combination have been reported (Huff et al., 2003). Few experiments on poultry have been conducted with bacteriophage as feed additives instead of dietary antibiotics. Therefore, this experiment was conducted to investigate the protective effects of anti-Salmonella enteritidis (anti-SE) bacteriophage on growth performance and against induced Salmonella enteritidis infection in broilers.
MATERIALS AND METHODS
Bacteriophage isolation and amplication
Anti-SE bacteriophage screening samples were collected from slaughter houses and sewage nearby disposal plants. Samples were centrifuged at 4,000 rpm for 10 min and the supernatants were filtered through a 0.45 μm filter. A mixture of 18 ml of filtered samples and log-phase Salmonella enteritidis in 2 ml of ten times strength Luria-Bertani (LB) medium (tryptone 10 g; yeast extract 5 g; NaCl 10 g; in a final volume of 1 L) was incubated at 37°C for 18 h and centrifuged at 4,000 rpm for 10 min. The supernatants were filtered through a 0.2 μm filter and then mixed with fresh log-phase Salmonella enteritidis and 3 ml of 0.7% agar. The fluid was poured into tryptic soy agar (TSA) plates and incubated overnight at 37°C. One clear plaque was selected and subsequently plated three times to purify the isolate, the purified plaque was suspended in SM buffer (20 mM Tris-HCl, 2 mM MgCl2 pH 7.0), and stored at 4°C until use.
The selected bacteriophage was shaken in Salmonella enteritidis cultured broth for amplification. After an aliquot of 1.5×1.010 cfu was centrifuged at 4,000 rpm, the pellet was re-suspended in 4 ml of a SM solution. The suspension was inoculated 7.5×107 pfu of the bacteriophage at an MOI (multiplicity of infection) of 0.005 and incubated at 37°C. After 20 min of incubation, this solution was inoculated on LB media in a flask and cultured at 37°C for 5 h. The solution was centrifuged at 4,000 rpm and then passed through a 0.2 μm filter. The bacteriophage was quantified by making serial dilutions and spreading on soft agar overlay plates. The final bacteriophage culture was collected and prepared as a powder. The plaque forming unit of 109 pfu/g was used as feed additives in the dietary treatments.
Salmonella enteritidis challenge culture
The Salmonella enteritidis strain used in current study provided by the Salmonella Genetic Stock Center (Calgary, AB, Canada). The isolate Salmonella enteritidis strain was grown overnight in LB broth (Difco, Franklin Lakes, USA) at 37°C. For the challenge trial, the Salmonella enteritidis culture was serially diluted in sterile phosphate buffered saline and marked clarify, and the cfu was determined to achieve the final concentration.
Experimental design, diet and animal managements
Three hundred and twenty 1-d-old male broilers (Ross 308), with an average body weight of 41.4±0.0 g were used for a 5 wk growth trial. Broilers were allotted into 8 blocks in a randomized complete block (RCB) design with 10 broilers per pen. The birds were raised on rice hull bedding in an environment controlled house at a research unit of Seoul National University experimental facility, South Korea. All broilers had free access to a feeder and an automatic waterer. The ambient temperature was maintained at 35°C for 2 d, 31°C in the first week and then gradually reduced to 22°C by the end of the experiment.
Two basal diets were formulated for the broilers during phase I and II of a 5 week feeding trial. Phase I diet was fed during 0 to 2 wks and phase II diet was provided thereafter until week 5. The basal diet contained 3,050 and 3,100 kcal of ME/kg and total lysine contents were 1.19 and 1.05% for the phase I and II, respectively. All other nutrients were formulated to meet or exceed the Korean Feeding Standard for Poultry (2007). The bacteriophage of 109 pfu/g used as feed additive was included at the expense of corn to the basal diet. The basal diets in this experiment was provided by manufacturer and contents in the diets were analyzed via the method of the AOAC (1995). The ingredient composition and calculated nutrients are shown in Table 1 and 2. Body weight (BW) and feed intake (FI) were recorded at the end of phase I and II periods to calculate the body weight gain and feed conversion ratio (FCR).
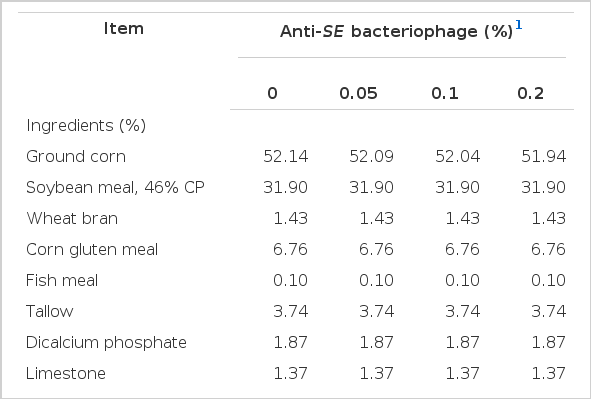
Composition of experimental diets (phase I, d 0 to 14); as-fed basis
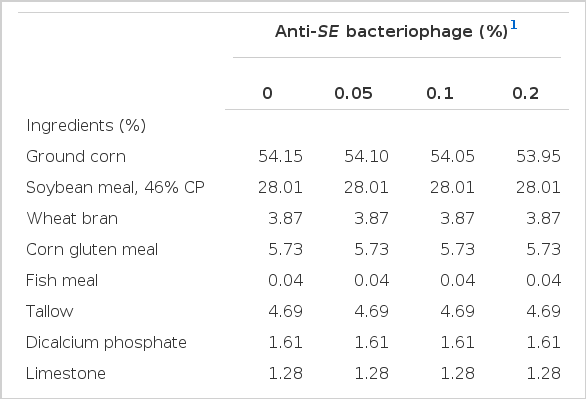
Composition of experimental diets (phase II, d 14 to 35); as-fed basis
Relative weights of tissue and blood sampling
After 5 week of growth trial, eight chickens from each treatment were randomly selected for sacrifice. The liver, spleen, abdominal fat, breast muscle and leg muscle were removed and weighed immediately for a relative weight calculation. Blood samples from 32 broilers that represented the 4 treatments were taken from the carotid artery. The samples were centrifuged for 5 min at 3,000 rpm at 18°C to collect serum and then carefully transferred to plastic tubes and stored at -20°C until analysis. Glutamic-oxaloacetic transaminase (GOT), glutamic-pyruvic transaminase (GPT), total cholesterol, HDL cholesterol and LDL cholesterol in blood were analyzed according to AOAC (1995).
Phage titer analysis
At the end of the 5 wk trial, 32 broilers (8 per treatment) were sacrificed to take sample of feed, feces and samples of breast and leg muscle to detect phage titer. Five grams of each samples were set in 10 ml SM buffer and homogenized for 1 min. 100 μl of the homogenizing sample was serially diluted (10-fold dilution) in SM buffer and plated on LB plate (Difco, BD science, USA). After 24 hrs, of incubation at 37°C the plaques were measured.
Mortality and cecal microflora after Salmonella enteritidis challenge
One hundred and sixty five-week-old male broilers (Ross 308), with average BW of 1,805.5±28.0 g were allotted from the previous treatments to the challenged by Salmonella enteritidis trial with 4 replicates and 10 birds per pen according to completely randomized design. The Salmonella enteritidis isolate was confirmed serotyped and the final concentration confirmed. For bacterial challenge, all broilers received 1×109 cfu of Salmonella enteritidis by oral inoculation and chicks were daily observed for mortality for 14 d. Feeding and environmental control were managed the same as in previous growth trial. Also, for the quantitative analysis of microflora, eight broilers were randomly selected from each treatment and sacrificed and the cecal content collected. For enumeration of bacterial populations, 1 g of cecal content was diluted in saline solution and re-suspended by vortex mixing, then serially diluted in saline solution. A volume of 100 μl volumes of each dilution was spread overlaid on the surface of an agar plate. This procedure was performed in duplicate over Salmonella shigella agar (supplemented 100 μl of chloramphenicol), MacConkey agar, Lactobacillus agar (Difco, BD science, USA). Plates were incubated at 37°C. After 24 h of incubation, each plate was spotted for cecal microflora count and examined for the presence of each bacterial growth. The protocol for the experiment was approved by the Institutional Animal Care and use Committee of the Seoul National University.
Statistical analysis
The experimental data were analyzed as a completely randomized design using the least significant difference (LSD) procedure of SAS (SAS Inst., Inc., Cary, NC, USA) and in randomized complete block design, the general linear model (GLM) procedure of SAS was performed. A pen was considered the experimental unit for data on growth performance and mortality. The individual broiler was used as the unit for data in the experiment on blood profiles and anatomy. Linear and quadratic effects for equally spaced treatments were assessed by orthogonal polynomial contrast to determine the effect of supplementation level of bacteriophage on all measurements. p<0.05 was considered statistically significant and p>0.05 to p<0.10 to be trended.
RESULTS AND DISCUSSION
Growth performance
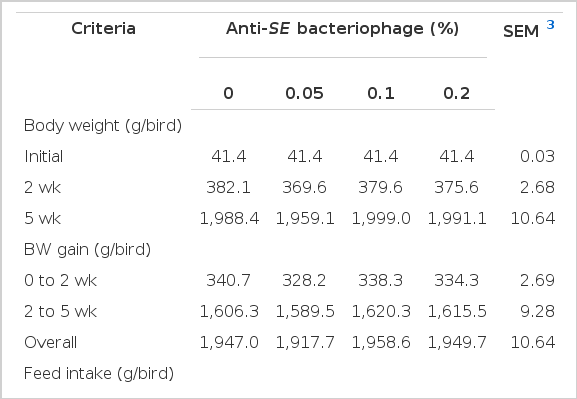
Growth performance of broilers fed the different levels of anti-SE bacteriophage in the diet during 5 weeks is shown in Table 3. Body weight (BW), BW gain, feed intake (FI) and feed conversion ratio (FCR) were not affected by dietary anti-SE bacteriophage supplementation at d 14 and d 35 of the experimental period (p>0.05). Although growth performance did not differ among treatments in the present study, it tended to have a positive relationship with 0.1% and 0.2% anti-SE bacteriophage. Additionally, based on the result of FI, anti-SE bacteriophage supplementation as a feed additive had no detrimental effect on feed consumption or broiler growth. As growth performance is one of the important parameters in poultry production, lower body weight equates with increase cost for broiler meat production. When the dietary anti-SE bacteriophage level was increased, FI was not affected by supplementation level of anti- bacteriophage. Other researches on the effect of bacteriophage (Barrow et al., 1998; Carlton, 1999; Huff et al., 2003) demonstrated that the pathogenic bacterium, Salmonella enteritidis, was clearly decreased by various methods of inoculation. When bacteriophage is utilized as a feed additive instead of antibiotics in animal feed then its inhibitive effect on harmful bacteria replication in the gastrointestinal tract can be expected (Yongsheng et al., 2008). Lim et al. (2010) demonstrated a high level of bacteriophage was excreted via feces when bacteriophage was supplemented to broilers diet. Moreover, their study concluded that broiler growth was improved by supplementation of bacteriophage due to the fact that bacteriophage inhibited the growth of Salmonella enteritidis in the gastrointestinal tract of broilers.
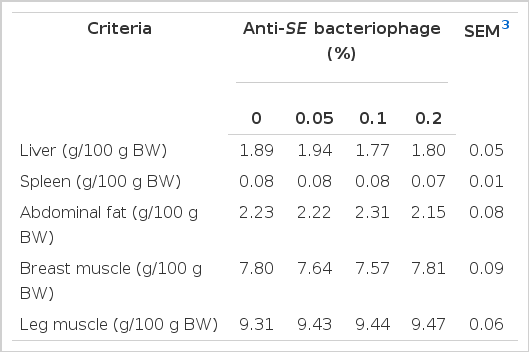
Relative weights of organ and muscle
The effects of the inclusion level of anti-SE bacteriophage on relative organ and muscle weight are presented in Table 4. There were no effects of supplementation of anti-SE bacteriophage on relative weight of organs at 35 d of the experiment (p>0.05). The relative weight of organs and muscle showed normal values compared to commercially produced broilers. However, the relative weights of leg muscle in the anti-SE bacteriophage treatments were numerically higher than that of the control treatment (p>0.05). Based on this result, the present study demonstrated a positive response in growth of broilers to bacteriophage supplementation rather than harmful effects such as growth retardation or toxicity. This result implies that the addition of anti-SE bacteriophage would not have a detrimental effect on poultry meat production. Additionally, broilers maintained a normal growth and morphological response in liver and spleen when anti-SE bacteriophage was supplemented. Merril (2008) reported that bacteriophage DNA was observed in the cells of Peyer’s patches of the gastrointestinal tract, peripheral white blood cells, and the cells of the liver and spleen after feeding of phage M13 DNA. Similarly, Molenaar et al. (2002) reported that M13 bacteriophage was found in liver, spleen, lung and kidney after oral administration of M13 bacteriophage by systemic circulation. Current and previous studies suggest that the accumulation of anti-SE bacteriophage in the liver and spleen is not harmful to animals.
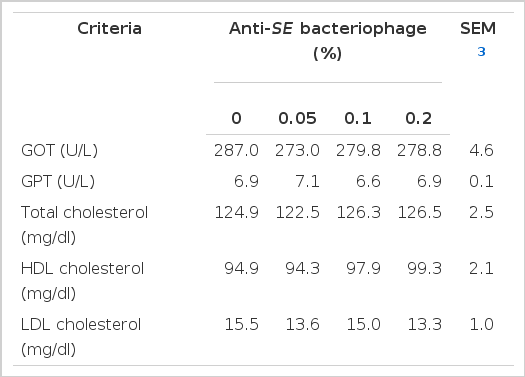
Blood profiles
Data for the blood profiles are presented in Table 5. In the presence of anti-SE bacteriophage supplemented in the diet, the responses of glutamic oxalo-acetic transaminase (GOT) and glutamic-pyruvic transaminase (GPT) were not different significantly (p>0.05) among treatments, even though the lower GOT concentration in blood was observed when broilers were fed anti-SE bacteriophage compared to control treatment. Serum GOT is a kind of cytosolic enzyme, consequently it can represent the status of liver damage when high level of GOT activity is observed in the blood. In the current study, the anti-SE bacteriophage supplementation in broilers diet did not induce damage to cells in the liver. There were no significance differences in total cholesterol, HDL (high density lipoprotein) and LDL (low density lipoprotein) between the anti-SE bacteriophage levels (p>0.05). However, supplementation of anti-SE bacteriophage in broilers diet tended to reduce the concentration of serum LDL cholesterol while numerically increased the serum HDL cholesterol concentration. Barrett-Connor and Suarez (1982) reported that serum GOT and HDL cholesterol have a negative relationship in humans. In the current study, the increased level of HDL cholesterol may be related to the reduction of GOT concentration. In this respect, supplementation of anti-SE bacteriophage as a feed additive would not damage hepatic or liver tissues of broilers and consequently it can be safely supplemented in broilers diets.
Phage titer
A greater phage titer in feed and feces was observed as the anti-SE bacteriophage level increased (Table 6). Moreover, phage titer was not detected from leg and breast muscle in any treatment. When broilers were fed anti-SE bacteriophage, anti-SE bacteriophage passed through the gastrointestinal tract and was excreted in the feces, consequently dietary anti-SE bacteriophage was not accumulated into the muscles. The inclusion of anti-SE bacteriophage in the diet could reduce intestinal Salmonella enteritidis level because high levels of anti-SE bacteriophage retarded their growth in the gastrointestinal tract. Additionally, the anti-SE bacteriophage can be used to reduce environmental contamination by Salmonella enteritidis when high levels of fecal anti-SE bacteriophage are excreted. The stability of bacteriophage under various conditions is an important parameter. Consequently, an evaluation of the stability of anti-SE bacteriophage is necessary prior to considering bacteriophage as an alternative to antibiotics (Mathur et al., 2003).

Mortality after Salmonella enteritidis challenge
Mortality of broilers fed different levels of anti-SE bacteriophage after Salmonella enteritidis challenge is shown in Table 7. Mortality after inoculation by Salmonella enteritidis was significantly decreased (p<0.05) as anti-SE bacteriophage level increased (p<0.05). It indicates that anti-SE bacteriophage parasitize Salmonella enteritidis bacteria resulting in a reduction of mortality. In general, infection by Salmonella enteritidis has been observed in both young and adult chickens. However, symptoms can be different for each broiler. Young chickens often develop a systemic disease with different mortalities (Suzuki, 1994; Duchet-Suchaux et al., 1995; Velge et al., 2005). In contrast, adult hens, after infection with Salmonella enteritidis, typically remained asymptomatic (Lister, 1988; Velge et al., 2005; Golden et al., 2008). These various responses in mortality of chickens challenged with Salmonella enteritidis might depend on Salmonella enteritidis phage types. Since the various strains of Salmonella enteritidis phage types are found in poultry, many researchers have observed that Salmonella enteritidis phage types are related to mortality in broilers. Dhillon et al. (1999) used six strains of Salmonella enteritidis to infect 1-d-old broilers by crop gavage and reported that the mortality range was from 0 to 23%. In addition, Barrow (1991) concluded that mortality ranged from 20 to 96% between six strains of Salmonella enteritidis.

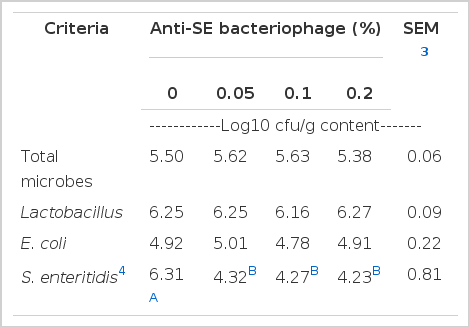
Cecal microflora after Salmonella enteritidis challenge
The quantitative total of the microbes, Lactobacillus, E. coli and Salmonella enteritidis in the cecal microflora were measured in the study of the effectiveness of anti-SE bacteriophage in broilers that were Salmonella enteritidis challenged (Table 8). The different inclusion levels to broiler’s diet did not alter the population of bacteria (total microbes, Lactobacillus, E. coli; p>0.05) after Salmonella enteritidis inoculation. However, the number of Salmonella enteritidis in cecal digsta from broilers fed all anti-SE bacteriophage diet was reduced compared to control in a linear response to supplementation levels (p<0.05). In general, bacteriophages have a high level of host specificity for certain species of bacteria (Ackermann et al., 1978; Joerger, 2003). Bacteriophage attaches to specific receptors of their bacterial host and they use the host's intracellular machinery to translate their own genetic code. Hence, anti-SE bacteriophage is harmless to the normal intestinal microflora (Oliveira et al., 2009). There are many studies regarding the utilization of bacteriophage to prevent infection with harmful bacteria (Huff et al., 2005; Sulakvelidze and Barrow, 2005; Sulakvelidze and Kutter, 2005). In the study reported by Borie (2007), inclusion of anti-SE bacteriophage by spray, drinking water and aerosol methods reduced the Salmonella enteritidis colonization in infected chickens. Data from the current study suggested that anti-SE bacteriophage showed therapeutic effects against Salmonella enteritidis colonization in the cecum.
CONCLUSION
Since the EU banned the use of antibiotics in animal feed, many countries have tried to prohibit the supplementation of antibiotics to farm animals and have searched for alternative materials particularly in the feed industry. As Salmonella enteritidis infection is still an important issue to maintain food safety, dietary bacteriophage has been suggested as a feed additive instead of antibiotics. This study demonstrated that dietary anti-SE bacteriophage from 0.05 to 0.2% of total diet can be useful to prevent Salmonella enteritidis infection and subsequently reduce mortality without any detrimental influences on broiler productivity. After consideration of their safety, stability and therapeutic efficacy in broiler production, the use of anti-SE bacteriophage in broiler diet can be an alternative feed additive instead of antibiotics.
Acknowledgements
This research was supported by CJ Cheiljedang Corporation, Seoul, Republic of Korea.