





INTRODUCTION
Cattle were some of the first animals to be domesticated by humans for agricultural purposes. About 10,000 years ago, the ancestors of cattle were tamed for the purpose of providing milk, meat, and leather to humans [1]. In 2006, a human genome sequencing center announced the sequence of the cattle genome [2]. Utilizing the genome-wide single nucleotide polymorphism panel for cattle allows for the use of quantitative trait loci (QTL) and prediction of the genetic merit of an animal without using phenotype and pedigree records [3]. Genetic studies using livestock have shown that most of the genetic variants associated with complex phenotypic traits are located outside the protein coding region [4].
Analysis of intracellular transcripts has revealed that 50% of the transcribed genomes were not aligned with known protein coding regions. Many of these have been proven to have protein coding potential [5]. Non-protein coding transcripts can potentially be noncoding RNA (ncRNA) [6]. Long noncoding RNA (lncRNA) is a molecule longer than 200 bp in a non-protein-coding transcript, or one longer than 2 kb with a coding potential of an amino acid sequence less than 100 bp [7]. LncRNA is a relatively new class of RNA molecules that are less well-characterized than microRNA (miRNA). It has been characterized only by some functional lncRNA and has been shown to regulate all levels of the gene-regulated expression pathway [8]. Previous studies have confirmed that lncRNA plays an important role in a variety of key biological processes, including translational control, RNA splicing, and chromatin modification [9]. In addition, functional studies of lncRNAs have shown that they play an important role in basic biological processes such as dose compensation, transcriptional regulation, and epigenetic regulation [10–12]. The number of studies on lncRNA has continued to increase and many databases have been constructed that include lncRNA data for domesticated animals [13]. At the time of the announcement, 12,103 pig lncRNAs, 8,923 chicken lncRNAs, and 8,250 cattle lncRNAs were included in the ALDB (domestic-animal lncRNA database) database [14].
This study was conducted to compare the expression of lncRNA in bulls, steers and cows as well as the characteristics of each sex. Samples from muscles tissues were used for each sex, and the expression of lncRNA by tissue was analyzed according to sex. We also analyzed QTL associated with lncRNA, which is significantly expressed.
MATERIALS AND METHODS
Sample preparation and RNA-seq snalysis
All analysis was conducted with data reprocessed from the law data from a previous study [15]. The animals and sample preparation were as follows: A total of nine (three bulls, three steers and three cows) Hanwoo cattle (Bos taurus coreanae) were used in this study. They were fed the same diet and managed at the same location, Hanwoo Experimental Station in National Institute of Animal Science, throughout the experiment. The average (±standard deviation) carcass weight of the cattle at slaughter was 430.2 (±40.66) kg. Immediately after slaughter, muscle tissues were separated and sampled. All of the tissue samples were immediately frozen using liquid nitrogen and stored at −80°C until analysis. All animal use, care, and experimental protocols for this experiment were reviewed and preapproved by the Institutional Animal Care and Use Committee of the National Institute of Animal Science (number 2010-042). Total RNAs of tissues were isolated using TRIzol (Invitrogen, Karlsruhe, Germany) and an RNeasy RNA purification kit with DNase treatment (Qiagen, Hilden, Germany). The messenger RNA (mRNA) was isolated from the total RNA using oligo-dT beads and was reverse transcribed into double-stranded cDNA fragments. Constructing and sequencing of the RNA-seq library for each sample was carried out based on the protocols of Illumina HiSeq2000 in order to generate 101 pair-end reads. The qualities of the RNA-seq reads from all of the tissues were checked using FastQC.
Analysis of lncRNA discovery
Filtering was conducted in order to remove the low-quality sequences. The filtered sequences were mapped to Bovine Taurus genome (bosTau6) using STAR v2.4.0b [16]. Expression levels were calculated using Cufflinks v2.2.1. Bovine gene information was used to measure expression levels [17]. Multi read correction and frag bias correct options were additionally used to increase the accuracy of expression measurement, and other options were used as defaults. The bovine lncRNA analysis was used to conduct mapping with reference to annotated bos taurus ensemble ID results. We refer to the lncRNA list of cattle identified in the study of Koufariotis et al [18]. We matched the ensemble ID of the transcripts we found and identified lncRNA in the muscle of Hanwoo (ensembl.org).
Statistical analysis
Differences in the expression level of each tissue were expressed by Heatmap using R package gplots (v3.0.1). LncRNA principal component analysis (PCA) plots were used to identify the differences between bulls, steers and cows muscle tissues using Mev (http://mev.tm4.org/). The DEGseq R package was used to identify differential expression of lncRNA between the bulls, steers and cows muscle tissues. Significant lncRNAs were identified using cut-off of: |fold difference| ≥1 and p-value ≤0.05. Sex-specific differentially expressed lncRNAs statistics analysis were using Prism 5 program (San Diego, CA, USA). The Venn diagram was used to identify the lncRNA assemblages that were extracted between the bulls, steers and cows. The Venn diagram was displayed using InteractiVenn (http://www.interactivenn.net/).
Quantitative trait locus analysis
QTL regions for comparative analysis with lncRNAs were identified from the cattle QTL Database (http://www.animalgenome.org/cgi-bin/QTLdb/BT/index). In these cattle, the QTL associated with the quality and productivity of the meat was selected. The position of the selected QTL was compared with the lncRNA expressed in sex.
Nearby gene analysis
The location of lncRNA was identified by ensembl biomart (ensembl.org/biomart) using transcript ID. The databases used included ensemble genes 92 cow genes (UMD3.1). We found for by nearby gene predicted to be affected by lncRNA. The position information of lncRNA in the genome and the gene position information of the bovine were matched using Python script. Transcription directions of lncRNA and nearby genes were used transcript information by ensembl (ensembl.org).
RESULTS AND DISCUSSION
Information of RNA sequencing
All analyses were conducted by reprocessing data from previous studies (Lee et al [15]). The RNA-seq raw data information is as follows: 34.2 Mb of raw readings were averaged on the muscle. More than 99.5% readings were retained after being filtered by quality control, and over 95.9% of these were mapped to the reference genome. The average length of the lncRNA was 866 bp, the minimum length was 257 bp, and the maximum length was 1,911 bp.
Expression patterns of lncRNA in Hanwoo cattle by sex
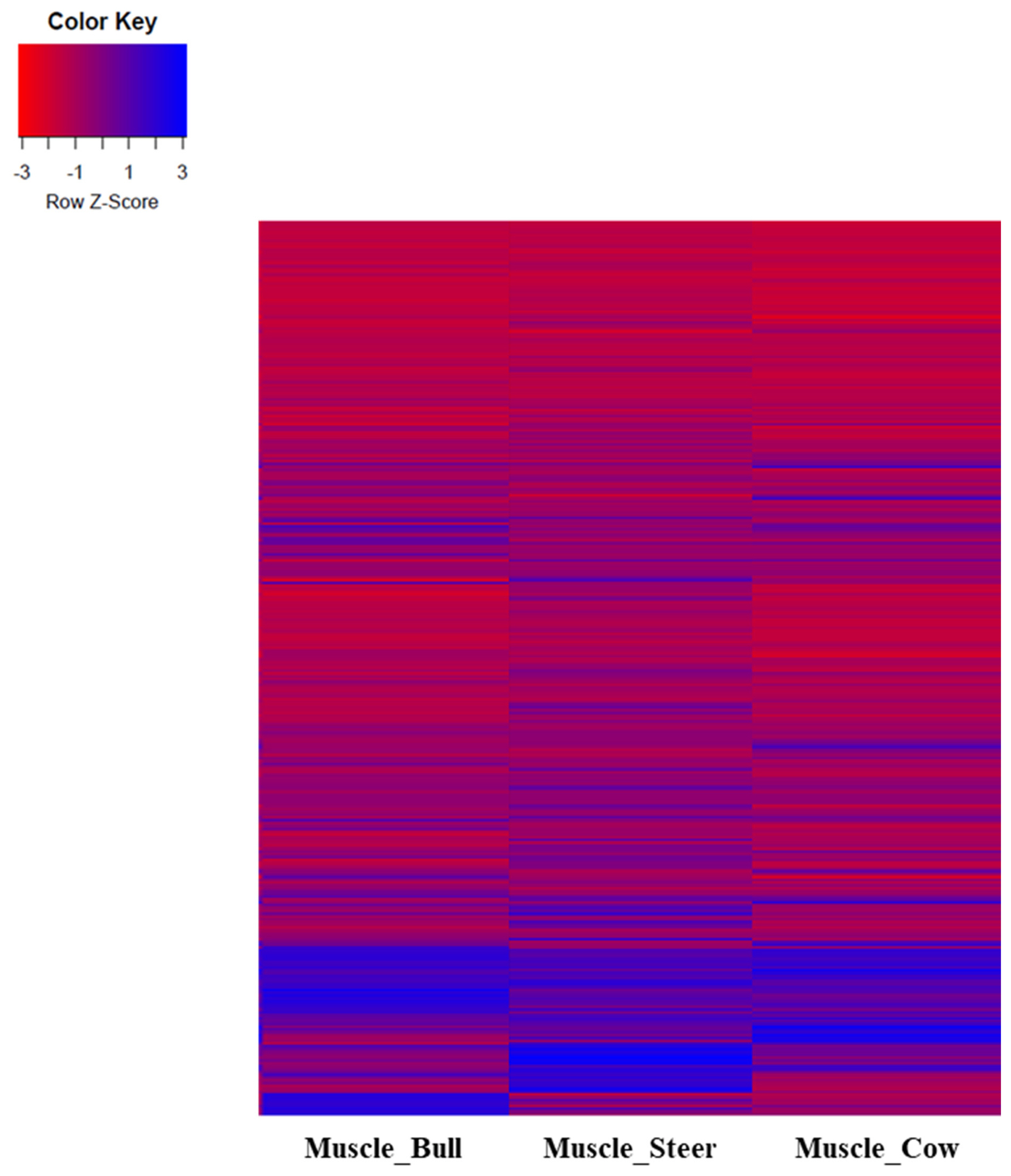
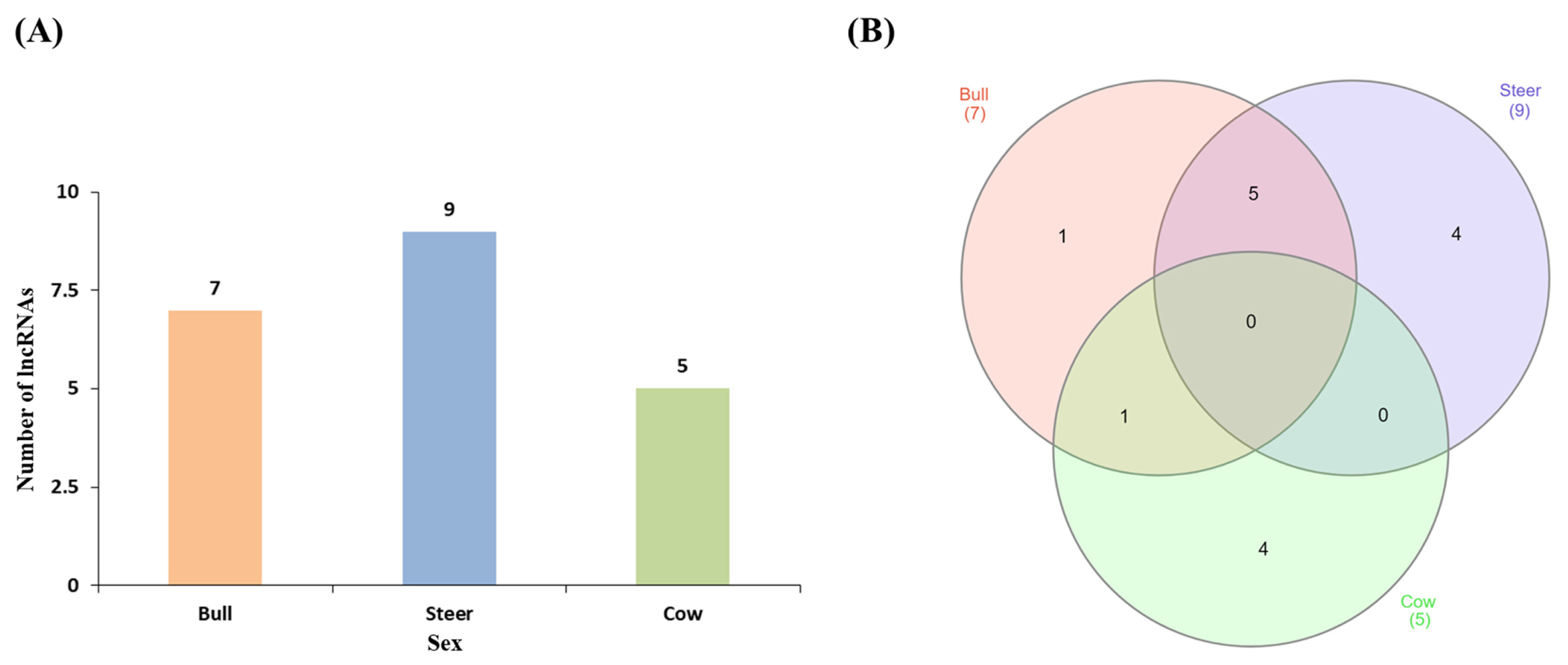
A hierarchical clustering analysis of the transcripts of sex transcripts in muscle tissue was performed in Hanwoo cattle. Clustering analysis of steer expressions did not show any significant differences (Figure 1). Clustering analysis using PCA analysis showed that the distances between the three groups differed by sex. The distance between the bull and the steer was relatively close to the distance to the cow (Figure 2). An analysis of the differential expression of lncRNA in each sex was conducted and found seven lncRNAs in bulls, nine lncRNAs in steers, and five lncRNAs in cows (Table 1, Figure 3A). The results of Venn diagram analysis for the found lncRNA revealed that 5 lncRNAs were found commonly in bulls and steers. We found one common lncRNA that was differentially expressed in bulls and cows. It was also identified that there was no overlapped lncRNA in the steers and cows (Figure 3B).
LncRNA-related bovine QTL analysis
Functional traits are defined as characteristics of animals that increase efficiency by reducing the cost of breeding [19]. Livestock species have long been selected to improve their characteristics that are associated with economic benefits. These traits are generally affected by genetic and environmental factors [20]. The detection of genetic markers closely linked to loci affecting quantitative traits (QTL) will provide a tool for the application of marker-assisted selection and is a prerequisite for the detailed molecular genetic analysis of traits [21]. Previous studies have identified 7188 lncRNAs from cows and have identified 2753 loci on chromosomal regions of livestock QTL related to muscle development. The function of QTL was also related to intramuscular fat, lean meat yield, longissimus muscle area, shear force [22].
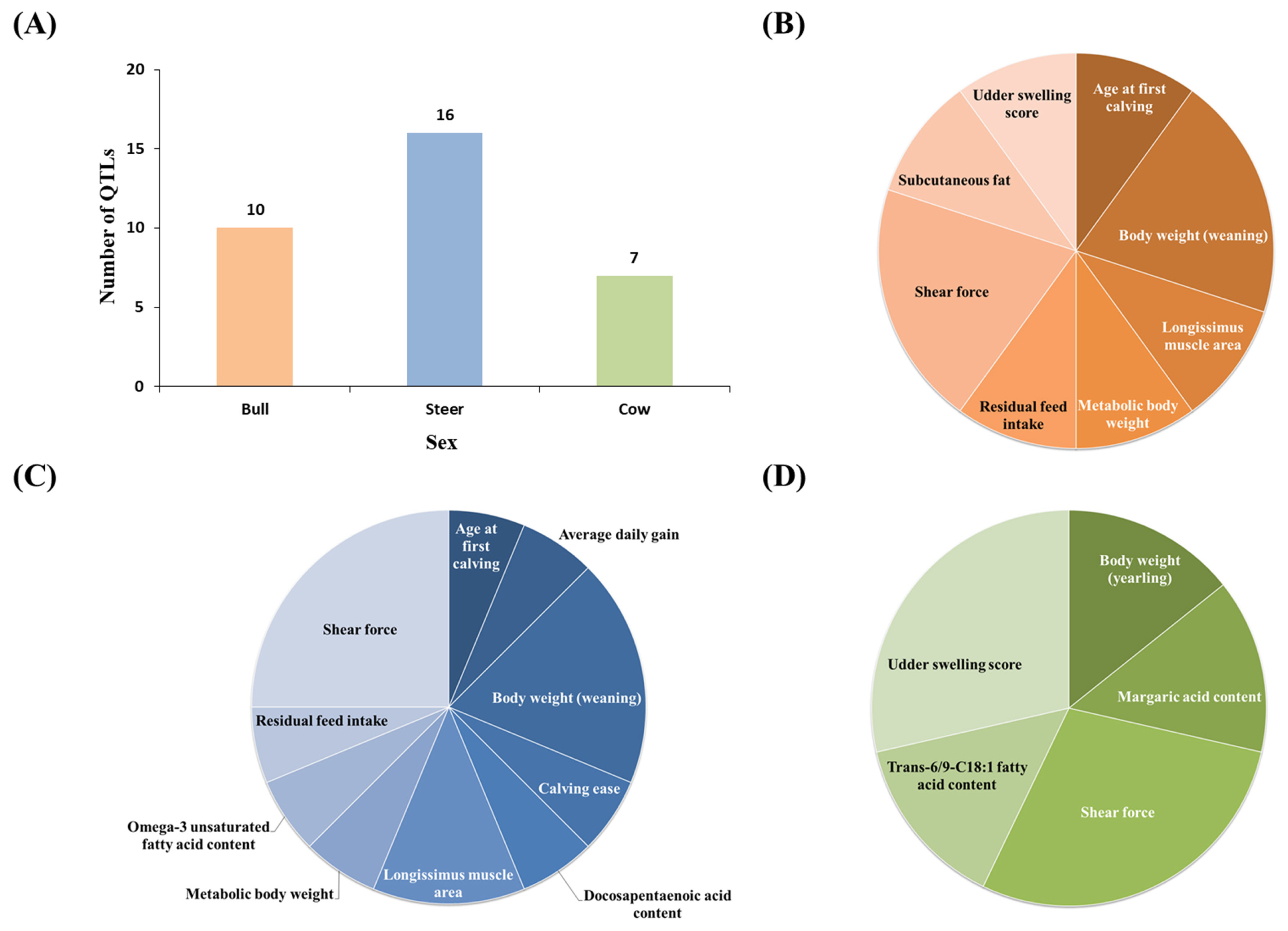
In this study, we conducted QTL analysis according to sex in Hanwoo muscle. QTL loci related to meat quality and productivity of cattle were selected with reference to a cattle QTL database. We identified 33 QTLs with overlapping loci with lncRNAs. Ten QTLs were identified in the bulls, 16 QTLs were identified in the steers, and seven QTLs were identified in the cows. In all sexes, lncRNA was found to be mainly associated with shear force QTL (Figure 4, Table 2).
LncRNA related genes identified
Most lncRNAs have lower conservation and expression levels than mRNA [23]. It has been reported that lncRNA can regulate the expression of adjacent protein coding genes through transcriptional activation and inhibition [24,25]. LncRNAs can be targeted by miRNAs and it has been reported that they can regulate the expression of mRNA [26,27]. The miRNAs consist of a length of about 22 nucleotides and function like silencing RNA and regulating gene expression after transcription [28,29]. In this study, three mRNAs were found to overlap with three lncRNAs. In addition, we identified and classified the transcriptional direction of lncRNA and mRNA (Table 3).
CONCLUSION
This study profiled lncRNA, which differentially expresses the difference of sex in Hanwoo cattle. We identified the expression patterns of transcripts in each sex by comparing the bull, steer and cow strains and identified 21 lncRNAs. We identified lncRNA related to meat quality and productivity according to sex in Hanwoo through QTL analysis. Many lncRNAs were found to overlap with QTL loci associated with shear force and body weight. We also identified adjacent genes that overlap with the three lncRNAs. It will be helpful to study the genetic characteristics of Hanwoo in the future.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Supplement
Supplement Print
Print







