Functional role of Forskolin and PD166285 in the development of denuded mouse oocytes
Article information

Abstract
Objective
cAMP and mature promoting factor (MPF) play critical roles during the maturation of mammalian oocytes. The aim of this study was to produce the offspring from denuded oocytes (DOs) in mice by regulating cAMP and MPF.
Methods
In this study, we used DOs at the germinal vesicle (GV) stage in mice and regulated levels of cAMP and MPF in DOs by adding Forskolin and PD166285 during in vitro maturation without follicle stimulating hormone and luteinizing hormone, respectively.
Results
Combined use of 50 μM Forskolin for 3 h and 2.5 μM PD166285 for additional 21 h enhanced the developmental competence of DOs, maturation rate of DOs was 76.71%± 4.11%, blastocyst rate was 18.33%±4.44% after parthenogenetic activation (PA). The DOs could successfully be fertilized with sperm in vitro, cleavage rate was 17.02%±5.82% and blastocyst rate was 5.65%±3.10%. Besides, 2-cell in vitro fertilization embryos from DOs produced 4 normal live offspring (4/34).
Conclusion
The results confirmed that the combination of Forskolin and PD166285 can induce DOs to complete meiosis process and produce normal offspring.
INTRODUCTION
Pluripotent stem cells have unlimited self-renewal capacity and plasticity to differentiate into all of the cell types even including sperm [1] and oocyte [2] under appropriate induction conditions. As a new kind of pluripotent stem cells reprogrammed from differentiated somatic cells, induced pluripotent stem (iPS) cells have already become a hot research topic in regenerative medicine. The related research about oocyte differentiation from iPS cells has opened up widespread applications in reproductive health and infertility in human beings and speeded up breeding process in domestic animals [3]. However, the study confirmed that pluripotent stem cells can differentiate into artificial oocytes with reproductive function only under in vivo conditions [4]. Under in vitro conditions, the oocytes derived from pluripotent stem cells still stay in the early stages of meiotic process and cannot complete meiosis [5,6]. In the process of oocyte differentiation from stem cells, stem cell-derived oocytes in development mode are similar to Dos in vitro, so the related development research of DOs will improve induction and application of stem cell-derived oocytes with reproductive function. Due to the lack of cumulus cells, stem cell-derived oocytes and DOs have poor ooplasmic quality and low development competence [7–10]. With the help of cumulus cells, stem cell-derived oocytes have high ooplasmic quality and development competence [11].
During oocyte development, cAMP and MPF play a critical role in ooplasmic quality and development competence respectively [12,13]. A high level of cAMP maintains oocyte arrest at the first meiotic prophase, under the condition of in vitro culture, transient increase of cAMP with Forskolin could induce meiotic resumption and increase maturation rate of DOs, but the DOs could not be performed in in vitro fertilization (IVF) [14]. PD166285 is a inhibitor of Myt1/Wee1B, Wee1B kinase is crucial to maintain G2 arrest through its inhibitory phosphorylation of cyclin-dependent kinases in cell cycles [15–19]. Further study demonstrates that the inhibition of Myt1/Wee1B pathway caused by PD166285 is necessary for the activation of MPF [19] which also regulates meiotic resumption. Therefore PD166285 may also be beneficial for the development potential of DOs. According to mechanisms during oocyte maturation, the combined application of Forskolin and PD166285 was carried out in a maturation process of DOs without an exogenous follicular development hormone in current study.
MATERIALS AND METHODS
Ethics statement
All animal procedures were approved by the Animal Health Care Committee of Anhui Agricultural University in the study.
Animals and chemicals
Eight-week-old female Institute of Cancer Research (ICR) mice were used in the study (Animal Research Center of Medical University of Anhui, Anhui, China). All chemicals and regents used in this study were purchased from Sigma Chemical Company (Saint Louis, MO, USA) unless otherwise noted. Stock solutions of Forskolin (20 mM) and PD166285 (5 mM) were dissolved in dimethyl sulfoxide (0.1%). Stock solutions were diluted with SAGE In-Vitro Maturation Media (IVM) (CooperSurgical, Trumbull, CT, USA) when used.
Collection and in vitro maturation
Female mice without superovulation were sacrificed by cervical dislocation, oocytes, including cumulus-oocyte complex and DOs, were released from antral follicles by puncturing with needles, surging into oocyte washing media. Morphologically intact natural DOs arrested in GV stage with same size (>75 μm in diameter) were collected, washed twice and then cultured in maturation media with Forskolin or PD166285. The culture dishes were covered with mineral oil in a humidified atmosphere containing 5% CO2, at 37°C. The maturation of oocytes was determined on the basis of first polar body (PB1) morphology after 24 h maturation.
The whole experiment was divided into two parts: DOs were cultured in Forskolin at different concentrations (10 μM, 50 μM, 100 μM) for 3 h, 12 h, or 24 h, then transferred into ordinary culture media for additional period (21 h, 12 h, or 0 h). DOs cultured in the oocytes maturation media was served as a control group.
In our experiment, female mice, injected 10 IU pregnant mare serum gonadotropin (PMSG) and followed by 10 IU of human chorionic gonadotropin (HCG) 48 h after PMSG injection, were used to collect in vivo matured oocytes 17 h after HCG injection. For in vivo developed embryos, female mice, were injected PMSG and HCG and then mated with male mice, were used to collect the embryos 24 h after HCG injection.
Parthenogenetic activation
Matured DOs were incubated in activating media, Ca2+-free CZB containing 10 mM SrCl2, for 2.5 h in a 5% CO2 humidified incubator at 37°C. After twice washes, the DOs were transferred into G-1 PLUS media (Vltrolife, Goteborg, Sweden). When developed into morulas, the embryos were transferred into G-2 PLUS media (Vltrolife, Sweden) for the further development. Embryos could be observed from the point when 20 h incubation was finished. The presumed embryos developed to 2-cell stages were defined as activated DOs. The development to morula and blastocyst stages were observed in about 2.5 d and 3.5 d after PA.
In vitro fertilization
Caudal epididymis from male ICR mice were washed twice in human tubal fluid (HTF) media (Vltrolife, Sweden), cut into pieces, dipped into the equilibrated HTF media for 10 min to release sperm, then the collected sperm were incubated in a tube for another 30 min at 37°C. Capacitated sperms in 3 to 5 μL supernatant were added on the periphery of the DOs collected previously in HTF media. After 6 h co-culture, presumed zygotes were transferred into G-1 PLUS media. The embryos were transferred into G-2 PLUS media for further development when developed into morulas. Zygotes in 2-cell stage would be observed after 24 h incubation. The development to morula and blastocyst stages were observed in about 2.5 d and 3.5 d after IVF.
Embryo transfer
Female mice at estrus were placed with male mice in the ratio of 2:1, checked copulatory plugs in the following morning after mating, determined as day 0.5 dpc, and then 0.5 dpc pseudopregnant female mice were anesthetized with 1% sodium pentothal. 2-cell IVF embryos were transferred into the oviducts of 0.5 dpc pseudopregnant female mice.
Immunofluorescence
Matured DOs or blastocysts were fixed with 4% paraformaldehyde in vitro for 30 min, washed 3 times, 5 min each in phosphate buffered saline and permeabilized in 1% Triton-X 100 for 15 min at room temperature. After washing 3 times, DOs were incubated in 1% bovine serum albumin for an hour, following by a whole night incubation in rabbit anti-α-Tubulin (1:500, Abcam) or CDX2 antibody (1:100, Abcam) at 4°C. DOs were kept in Cyanine Dyes 3-conjugated goat-rabbit immunoglobulin G (1:200, Abcam) for an hour, washed thoroughly, then transferred into 1 μg/mL DAPI for 10 min. Coverslips were mounted when DOs were transferred into glycol on the glass slides. DOs were observed through the immunofluorescence-microscope (Olympus, Tokyo, Japan).
Statistical analysis
There were at least three replicates for each treatment. Data were analyzed with analysis of variance, a least significant difference (LSD) multiple comparison test was used to evaluate differences using SPSS17.0 (Chicago, IL, USA). Statistical significance was considered to be at p<0.05. Data are presented as mean±standard error of the mean.
RESULTS
Morphological characteristics
In current study, GV stage DOs were cultured in maturation media supplemented with Forskolin and PD166285. GV stage DOs had normal size and shape and uniform cytoplasm and exhibited an intact GV with distinct nuclear membrane (Figure 1A). Matured DOs could reach morphological maturation and extruded PB1 24 h after IVMregardless of adding Forskolin or PD166285 in maturation media or not, and had no significant difference with in vivo mature oocytes in appearance (Figure 1B, 1C).

Morphological characteristics of mouse denuded oocytes (DOs). (A) DOs at the germinal vesicle (GV) stage. (B) Matured DOs with PB1 24 h after the treatment with 50 μM Forskolin for 3 h and 2.5 μM PD166285 for additional 21 h. (C) In vivo matured DOs.
Effect of Forskolin and PD166285 on maturation
Forskolin has a cell-intrinsic function in regulating oocyte maturation by activating adenylate cyclase to increase cAMP levels. In the IVM process, Forskolin had no significant effect on the survival of mouse DOs (Figure 2A). In order to investigate the effect of Forskolin on the maturation in mouse DOs, DOs were cultured in maturation media supplemented with Forskolin. We cultured GV stage DOs in maturation media containing 10, 50, and 100 μM Forskolin for 3 h, and then transferred the DOs in maturation media without Forskolin for additional 21 h, maturation rates of DOs were 53.64%± 1.42%, 50.73%±1.48%, and 54.68%±1.41% after 24 h incubation respectively. GV stage DOs in maturation media containing 10, 50, and 100 μM Forskolin were cultured for 12 h, and then additional 12 h, maturation rates were 49.39%±3.68%, 73.58% ±1.39%, and 49.85%±2.54%. When GV stage DOs in maturation media containing 10, 50, 100 μM Forskolin were cultured for 24 h, maturation rates were 61.88%±2.14%, 48.60%±1.68%, and 56.23%±2.02% (Figure 2B). In addition, when cultured in maturation media containing Forskolin over 20 h, GV stage DOs spontaneously began to enter into meiosis and take on morphological maturation to extrude PB1.
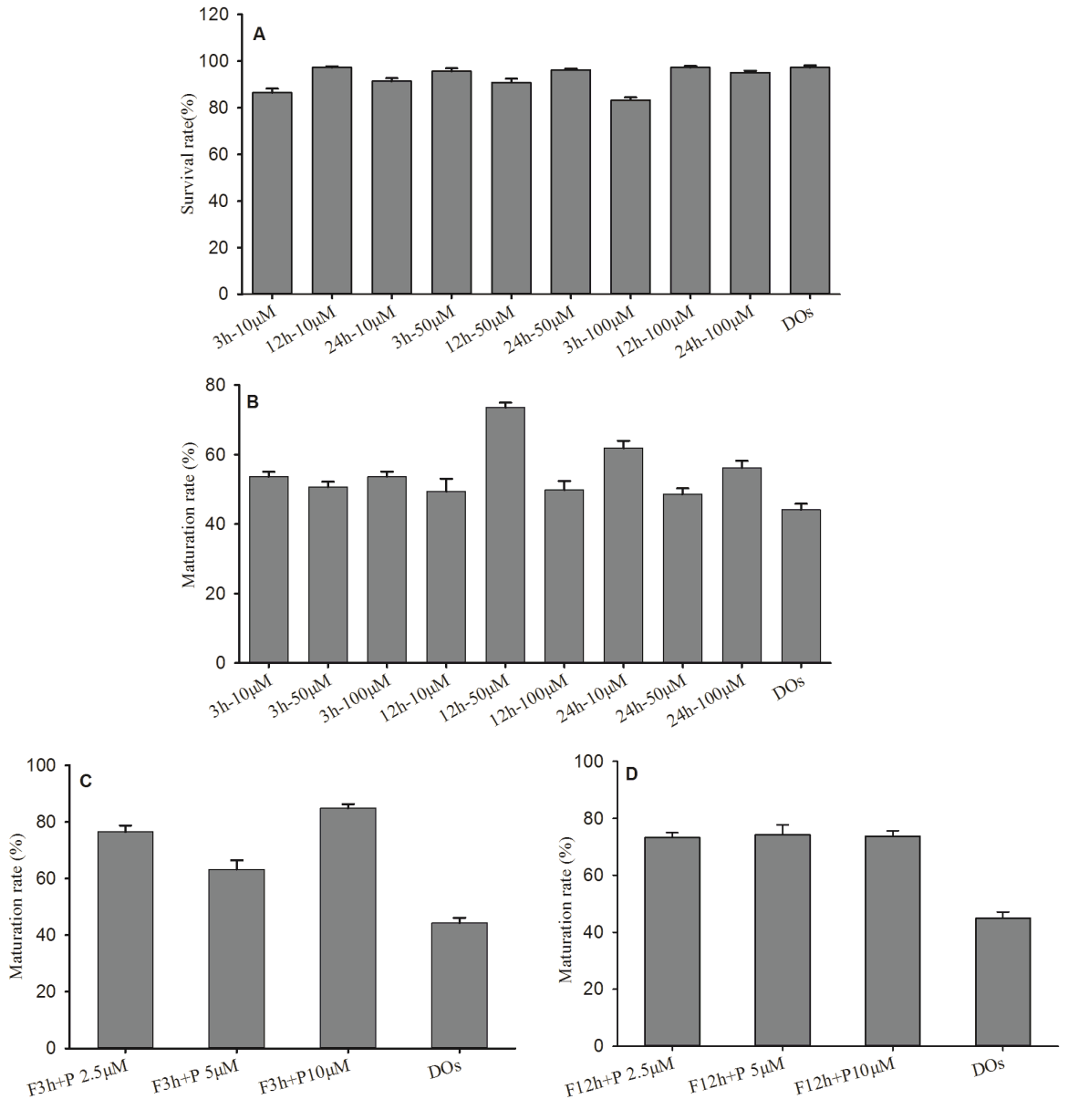
Forskolin and PD166285 promoted in vitro maturation of mouse denuded oocytes (DOs). (A) Effect of Forskolin on the survival rate of mouse DOs in vitro. (B) Effect of Forskolin on the maturation rate of mouse DOs in vitro. (C) Maturation rate of mouse DOs with Forskolin treatment for 3 h and PD166285 for additional 21 h. (D) Maturation rate of mouse DOs with Forskolin treatment for 12 h and PD166285 for additional 12 h.
PD166285 was described as a Wee1B kinase inhibitor to elevate MPF levels in oocytes [15]. We cultured GV stage DOs in maturation media containing 50 μM Forskolin for 3 h, and then transferred the DOs in maturation media with 2.5, 5, 10 μM PD166285 for additional 21 h, maturation rates were 76.55%±2.28%, 63.23%±13.26%, and 84.84%±1.44% (Figure 2C). In addition, the DOs in maturation media containing 50 μM Forskolin were cultured for 12 h, and then transferred in maturation media with 2.5, 5, and 10 μM PD166285 for additional 12 h, maturation rates were 73.27%±1.75%, 74.25%± 3.52%, and 73.73%±1.97% (Figure 2D). No significant difference was recognized in each experiment, this indicated that Forskolin and PD166285 did not significantly affect oocyte development and maturation in appearance.
Effect of Forskolin and PD166285 on parthenogenetic activation
We found that only adding Forskolin in the medium could facilitate the maturation in mouse DOs, but the matured DOs had a low development potential and failed to develop into blastocyst stage after PA, so we added PD166285 in the medium to regulate the maturation of DOs. During IVM of DOs, we cultured GV stage DOs in maturation media containing 50 μM Forskolin for 3 h, and then transferred the DOs in maturation media containing PD166285 for additional 21 h, the matured DOs in Forskolin and PD166285 groups could be activated, and activation rates were 42.66%±2.15% for 2.5 μM PD166285, 24.81%±2.66% for 5 μM PD166285 and 45.83%± 3.07% for 10 μM PD166285 (Figure 3A). Under maturation condition of 50 μM Forskolin for 12 h and PD166285 for additional 12 h, activation rates were 69.76%±3.03% for 2.5 μM PD166285, 45.85%±1.94% for 5 μM PD166285 and 39.93%± 2.79% for 10 μM PD166285 (Figure 3B). In addition, the matured DOs in Forskolin for 3 h and PD166285 for additional 21 h groups could develop into blastocysts after PA, and blastocyst rates were 18.11%±2.73% for 2.5 μM PD166285, 9.15% ±3.14% for 5 μM PD166285 and 2.94%±0.28% for 10 μM PD 166285 (Figure 3C). But the matured DOs could grow into blastocysts only from 2.5 μM PD166285 groups under maturation condition of Forskolin for 12 h and PD166285 for additional 12 h, and blastocyst rate were 2.5%±0.24% (Figure 3D).
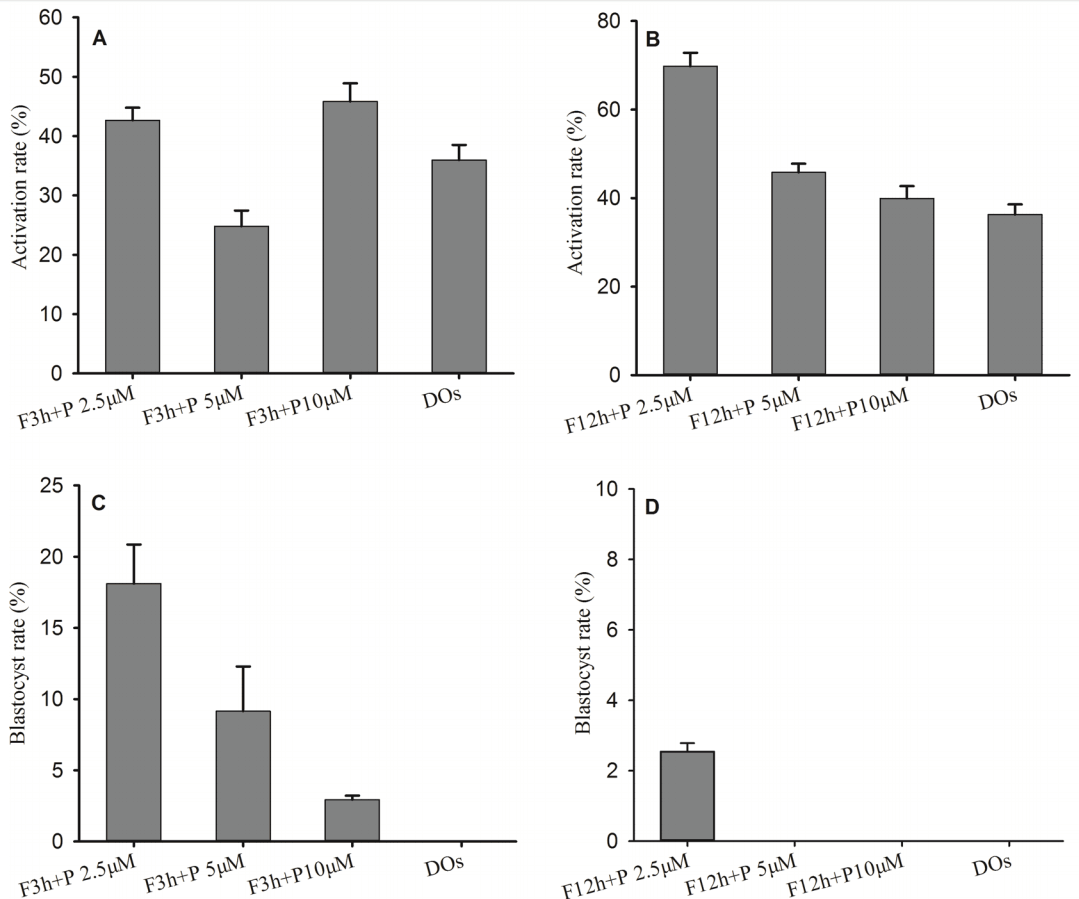
Forskolin and PD166285 promoted embryo development from mouse denuded oocytes (DOs) after parthenogenetic activation. (A) Activation rate with Forskolin treatment for 3 h and PD166285 for additional 21 h. (B) Activation rate with Forskolin treatment for 12 h and PD166285 for additional 12 h. (C) Blastocyst rate with Forskolin treatment for 12 h and PD166285 for additional 12 h. (D) Blastocyst rate with Forskolin treatment for 12 h and PD166285 for additional 12 h.
Effect of Forskolin and PD166285 on IVF
Under the condition of only adding Forskolin in maturation media or Forskolin for 12 h and PD166285 for additional 12 h, the matured DOs could not become competent to be fertilized sperm in vitro. The matured DOs could be fertilized sperm to develop into embryos only under the condition of 50 μM PD166285 for 3 h and 2.5 μM PD166285 for additional 21 h. The embryos in an early stage had a highly symmetrical appearance and well developed into blastocysts (Figure 4A–4D), the cleavage rate was 17.02%±5.82% and blastocyst rate was 5.65%±3.11%. To further study the development potential in vivo of matured DOs treated with 50 μM Forskolin and 2.5 μM PD166285, 34 fertilized embryos in 2-cell stage from matured DOs were transferred into fallopian tubes of surrogate mice by the transplantation of tubal umbrella. Four normal live pups (4/34) from mouse DOs were produced and had normal fertility (Figure 4E).
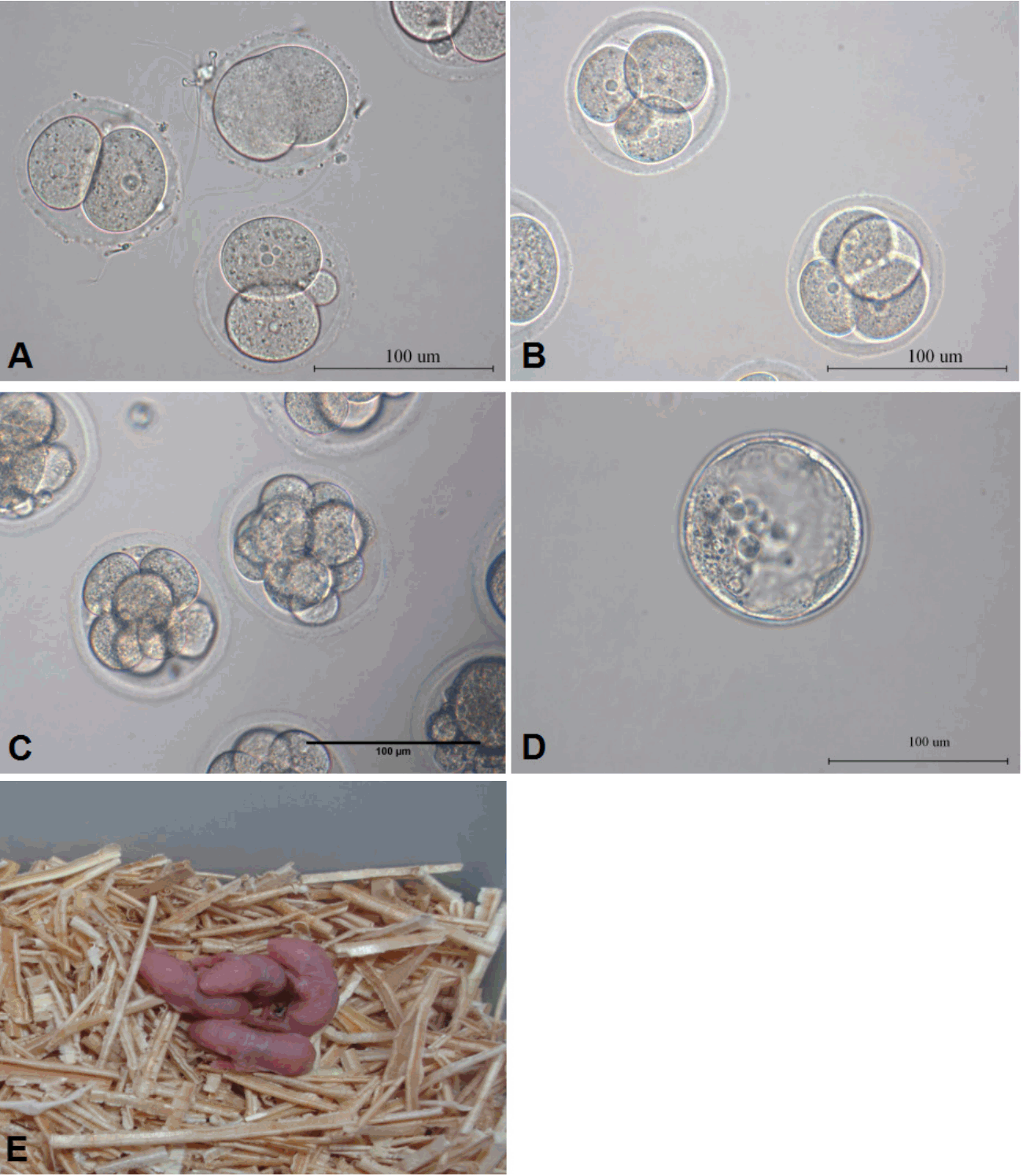
Forskolin and PD166285 promoted embryo development from mouse denuded oocytes (DOs) after in vitro fertilization (IVF). (A) 2-cell embryos from mouse DOs after IVF. (B) 4-cell embryos from mouse DOs after IVF. (C) 8-cell embryos from mouse DOs after IVF. (D) The blastocyst from mouse DOs after IVF. (E) Newborn live offspring from mouse DOs after IVF.
Effect of Forskolin and PD166285 on spindle assembly
The formation of normal meiotic spindles during oocyte maturation is very important to developmental potential. GV stage DOs in maturation media only containing Forskolin or without Forskolin were cultured, and results showed that matured DOs (71/82) had loose chromosomes, irregular microtubules, asymmetrical spindle, and no obvious poles. GV stage DOs in maturation media containing 50 μM Forskolin for 3 h and 2.5 μM PD166285 for additional 21 h had matured DOs (83/94) that showed a similar spindle image with in vivo natural matured oocytes (92/102), mainly exhibited a symmetrical spindle, aggregated chromosomes at equatorial plate and consistent appearance of microtubules (Figure 5), so the condition of 50 μM Forskolin for 3 h and 2.5 μM PD166285 for additional 21 h is of great benefit to spindle assembly of mature DOs.

Forskolin and PD166285 regulated spindle organization and chromosome alignment. Control showed in vivo-derived natural matured oocyte, immunofluorescence localization of spindles (red) and chromosome (blue) in mouse oocytes. Scale bar = 50 μm.
Effect of Forskolin and PD166285 on blastocyst quality
Blastocysts quality was further evaluated by cell counting. During IVM of DOs treated with Forskolin for 3 h and PD166285 for additional 21 h, the blastocysts from DOs by PA and IVF had a normal image and development potential. Blastocyst count showed that the total cell number in blastocysts from the DOs was 39.83±3.65 (Blastocyst number 51) and 39.67± 13.68 (Blastocyst number 72) after PA and IVF respectively, in vivo-derived blastocysts about 41.67±9.53. The ratio of inner cell mass to total cell number was 32.39%±5.20% and 25.06% ±8.75% for PA and IVF blastocysts from the DOs respectively, in vivo-derived blastocysts about 31.04%±4.07% by immunofluorescence staining (Figure 6). The difference between the groups of DOs and in vivo-derived blastocysts was not significant.
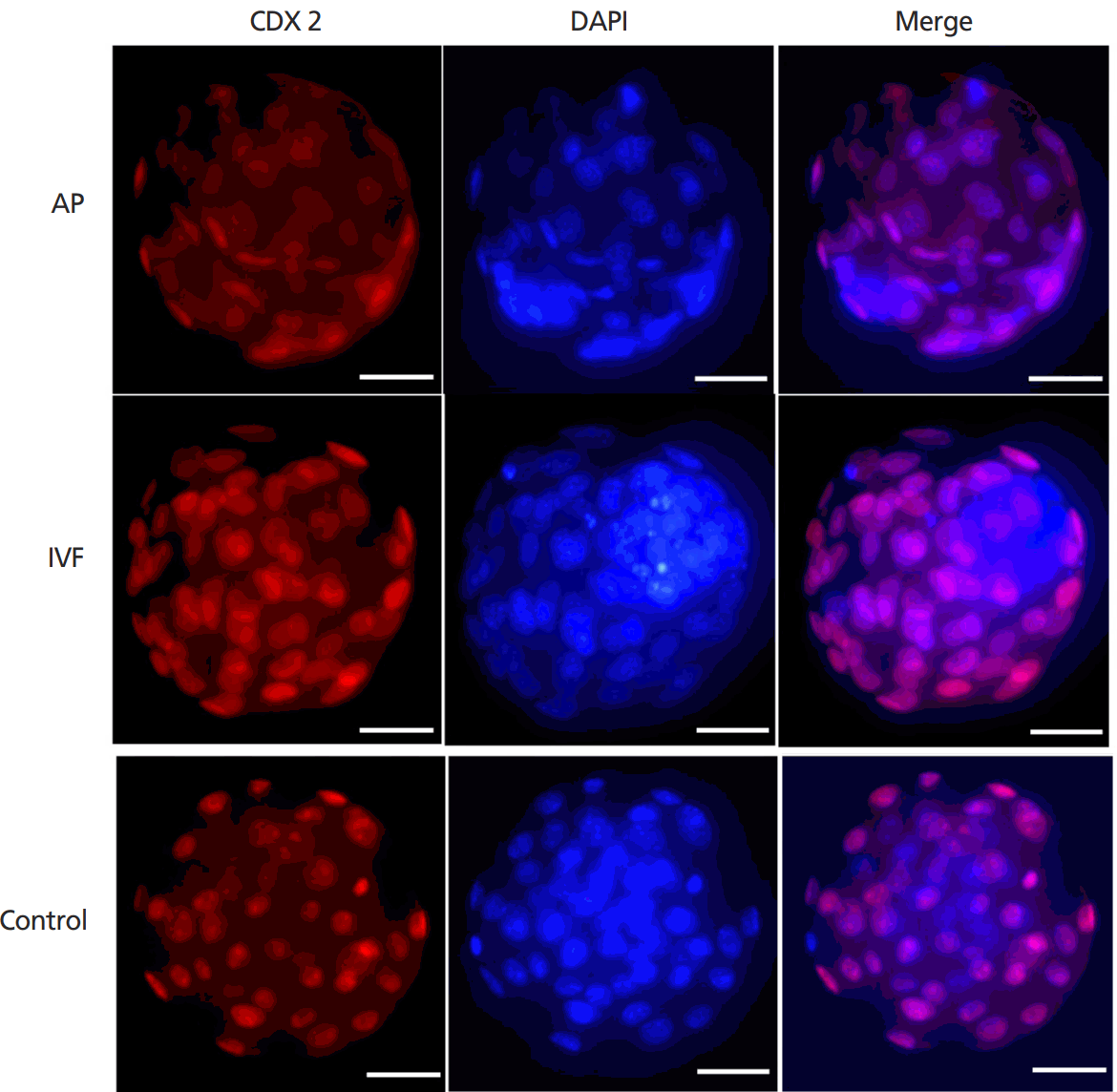
CDX2 immunofluorescence staining of blastocysts generated from mouse denuded oocytes (DOs). Control showed in vivo-derived natural blastocysts. Scar bar = 50 μm.
DISCUSSION
cAMP as a meiosis inhibitor regulates oocyte cytoplasmic maturation and enhances the quality of the cytoplasm through gap junctions of the surrounding cumulus cells and oocytes [8,20]. It was found that the short-term cAMP effect (cAMP-pulsing) induced by adding Forskolin during IVM of DOs could improve maturation efficiency, but the developmental potential of DOs was not improved [14]. In this study, we simulated the changes of cAMP and MPF during oocyte maturation, took advantage of the cAMP agonist Forskolin and MPF agonist PD166285 to regulate DOs during IVM, and established a simple maturation system in mouse DOs by regulating the level of cAMP and MPF in vitro. By adding 50 μM Forskolin for 3 h and 2.5 M PD166285 for additional 21 h during IVM, mouse DOs had a high development potential after PA and IVF embryos could develop to blastocyst in vitro and produce healthy newborn offspring in vivo. The high level of cAMP plays an important role to induce meiotic division arrest and cytoplasmic maturation and activate the cAMP/PKA pathway during oocyte maturation [21]. In current study, we found that mouse DOs spontaneously extruded PB1 more than 20 h after the use of Forskolin during IVM. It shows that cAMP has an inhibitory timing effect on mouse DOs and other key factors also take part in the maturation and development with the cAMP. During the development of oocytes, the level of intracellular cAMP was accompanied by the formation of microtubules and the maturation of nuclei, and the functional MPF was activated to promote the first meiotic division of oocytes [22,23]. The experimental results also confirmed that mouse DOs treated only with Forskolin had no development potential in either parthenogenetic development or the fertilization with sperm. In fact, cAMP has a limited regulatory capacity in the process of meiosis and MPF is also involved in the regulation of meiosis [24,25]. It is particularly important that MPF plays a key role in the process of oocyte from the first meiotic division to the second meiotic division [26]. In the oocytes of the first meiotic prophase, MPF is present in the form of low activity pre-MPF [27]. After the resumption of meiosis, the level of MPF increases rapidly, reaching the first peak in spindle formation, and promotes oocyte maturation [28]. In this study, the mature mouse DOs had IVF ability after treatment with transient Forskolin and then MPF agonist PD166285, and fertilized embryos from mouse DOs, like normal embryos, could develop into normal live offspring in vitro. It showed that mouse DOs had strong development potential and became functional oocytes after artificial regulation according to the law of oocyte development, and the molecular mechanism and basic process of oocyte meiosis were also further confirmed by this model.
In current study, we established a simple and efficient culture system for mouse DOs during IVM and laid an important foundation for producing artificial oocytes with reproduction function and focusing on meiosis mechanism by simulating in vivo cAMP-MPF regulation in oocytes.
ACKNOWLEDGMENTS
This work is supported by grants from by National Natural Science Foundation of China (31372309) and National High-Tech R&D Program (2011AA100307).
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.