





INTRODUCTION
Intrauterine growth retardation (IUGR) is defined as a fetal weight below the 10th percentile and uterine growth below the 2.5 percentile for a given gestational age [1]. The mammalian embryo/fetus with IUGR usually shows retarded development during gestation and organ systems malfunction later in life. The naturally-occurring IUGR pig was recognized as an ideal model for the study of human IUGR [2]. Based on our previous study, newborn IUGR piglets had fatty hepatic infiltrates and cytoplasmic vacuolization in the liver [3]. Investigation of the hepatic proteome also suggested that the IUGR fetus had a lower activity of lipoprotein lipase (LPL) [4]. Abnormal lipid metabolism in the liver has proven to be closely related to the development of metabolic syndrome (MetS) [5], which has been widely observed in adult IUGR offspring exhibiting catch-up growth [6]. Although significant metabolic abnormalities have been observed in adult IUGR rats exhibiting catch-up growth [7], very few studies have been conducted that examine the hepatic lipid metabolism of adult IUGR pigs.
As a common B vitamin in feed, choline serves as a methyl group donor and a precursor of phosphatidylcholine (PC), which is involved in very low-density lipoprotein (VLDL) assembly in the liver. Although the improvement in growth performance and the decrease in liver lipid content as a result of choline supplementation in poultry rations are well established, studies of choline in pigs are limited and the results are inconsistent [8]. The discrepancy regarding cholineŌĆÖs effects on growth and lipid metabolism may be related to species differences, the level of choline supplementation, or the length of the experiment period. As the choline oxidase activity in pig livers is relatively lower compared with broilers [9], we speculated that choline or its metabolites might take more time to show significant effects in pigs. It has been reported that IUGR pigs have a significantly decreased serum choline content compared with normal pigs [10], suggesting that more choline in the diet might be necessary for IUGR pigs to maintain normal lipid metabolism, including PC synthesis. In this study, we investigated the effect of long-term choline supplementation on lipid metabolism and hepatic gene expression in IUGR pigs.
MATERIALS AND METHODS
Animals and experimental design
Pigs from Large White├ŚLandrace sows mated to Landrace boars were used in this study. During the entire pregnancy period, sows were fed a gestation diet (2 kg/d, meeting the NRC [11] recommendations for nutrient intake) and had free access to water. Piglets with a birth weight within 1 standard deviation (SD) of the mean birth weight were identified as normal birth weight (NBW), and piglets with a birth weight more than ŌłÆ2 SD below the mean birth weight were defined as IUGR, as previously described [1]. At the time of parturition (d 114┬▒1 of gestation), a total of eight pairs of male piglets (eight NBW pigs: 1.51 kg┬▒0.11; eight IUGR pigs: 0.93 kg┬▒0.10) were selected from eight sows of equal parity (fourth parity) and a similar body weight (BW). One NBW pig and one IUGR pig were selected from each litter. The NBW and IUGR piglets were cross-fostered within 48 h after birth by two four-parity sows (eight piglets for each sow). The two sows were fed with a lactation diet that met NRC [11] requirements. Piglets were surgically castrated at 7 d.
After weaning at 23 d of age, all piglets were allotted to one of four experimental treatments, which were arranged in a 2├Ś2 factorial design consisting of intrauterine growth type (IUGR or NBW) and diet (normal or high choline) (n = 4). The basal diet with a normal quantity of choline was formulated to meet or exceed NRC [11] nutrient requirements for post-weaning (d 23 to 73), growing (d 74 to 120), and finishing periods (d 121 to 200) (Table 1). The high-choline diet was produced from the basal diet by displacing the same quantity of corn with choline (50% choline chloride; Shandong Enbei Group Co., Ltd, Jinan, China). The total choline content in the high-choline diet was 2 times greater than the basal diet (Table 2). The dietary choline chloride concentration was determined using the Reineckate method [12], and choline concentration was measured via chromatographyd. Pigs were reared in individual pens (2.0 m├Ś4.0 m) equipped with one nipple drinker and a one dry feeder on fully slatted concrete floors. Pigs had free access to diets and water. During the experimental period, room temperature was controlled by ventilation fans and a heater. Room temperature, feeders, drinkers, and animal status were checked at least twice a day.
Pigs were slaughtered at 200 d of age after an overnight fast, by electro-stunning followed by exsanguination at a local commercial slaughterhouse. All experimental procedures used in the study were consistent with the Guide for the Care and Use of Laboratory Animals prepared by the Institutional Animal Care and Use Committee, Nanjing Agriculture University, China.
Data and sample collection
The BW was measured at the end of each period and the average daily gain (ADG) was calculated. At these same ages, blood samples (5 mL) were collected from all pigs via venipuncture after a 12 h overnight fast. Blood was allowed to clot at room temperature and was stored at 4┬░C for 4 h prior to harvesting serum via centrifugation (1,500├Śg for 15 min at 4┬░C). Serum samples were stored at ŌłÆ20┬░C for subsequent analyses.
All tissue samples were isolated and harvested within 15 min after death. Liver samples were taken from the left lateral lobe. The samples for enzyme analyses and mRNA extraction were flash-frozen in liquid nitrogen and stored at ŌłÆ80┬░C until analyzed.
Serum measurement
The serum free fatty acid (FFA) was determined by the colorimetric method based on the formation of FFA-Cu soaps [13]. The alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities were determined by ReitmanŌĆōFrankel methods [14]. The triglyceride concentration was estimated by glycednphosphat oxidase-phenol and 4-aminophenazone method and the total cholesterol was measured by the cholesterol oxidase coupled to phenol and 4-aminophenazone method [15]. The determination of high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), LPL activity and hepatic lipase (HL) activity were carried out according to the instruction manual provided by the manufacturer. All parameters were analyzed in duplicate by colorimetry using commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Enzyme analyses
The liver tissues were homogenized in a buffer containing 0.25 M sucrose, 0.1 M phosphate, 1 mM ethylenediaminetetraacetic acid (EDTA), and 10 mM ╬▓-mercaptoethanol (pH 7.0). The homogenates were centrifuged at 105,000 g for 1 h at 4┬░C and the supernatants were collected and used for enzyme assays.
Spectrophotometric assays were used to determine the activity of glucose 6-phosphate dehydrogenase (G6PDH) [16], isocitrate dehydrogenase (ICDH) [17], malate dehydrogenase (MDH) [18], and malic enzyme (ME) [19]. All assays were run in a reaction buffer with a total volume of 1 mL in triplicate. For G6PDH, the buffer consisted of 50 mM Tris (pH 7.6 Tris), 0.15 mM NADP+, 20 mM MnCl2, 1 mM glucose-6-phosphate and 0.1 mL supernatant. For ICDH, the buffer consisted of 50 mM Tris (pH 3.4 Tris), 0.15 mM NADP+, 1.5 mM MnCl2, 0.34 mM EDTA, 0.03% glutin, 1.5 mM D, LŌłÆ isocitrate and 0.1 mL supernatant. For ME, the buffer consisted of 50 mM Tris (pH 7.4 Tris), 0.15 mM NADP+, 1 mM MnCl2, 3 mM D, L-malate and 0.1 mL supernatant. The buffer for MDH was obtained from the commercial kit (Nanjing Jiancheng Bioengineering Institute, China).
All assays were conducted within the range of linearity at 340 nm and 37┬░C with respect to the relation between enzyme activity and time, and enzyme activities were expressed on a per milligram of protein basis using bicinchoninic acid protein assay kit (Nanjing Jiancheng Bioengineering Institute, China).
Total RNA extraction and quantitative real time-polymerase chain reaction
Total mRNA was extracted from liver samples using Trizol reagent (Invitrogen, Shanghai, China), according to the manufacturerŌĆÖs protocols, with minor modifications. Briefly, 200 mg of liver sample was added to 1 mL Trizol reagent and homogenized with a homogenizer (Pro 200, Pro Scientific Inc., Oxford, CT, USA) at full speed for 8 min (12 times for 30 s, with a 10 s pause between repetitions). All RNA was treated with RNase-free DNase (Takara Biotechnology Co. Ltd., Dalian, China) to remove any trace of genomic DNA. The extracted RNA was dissolved in diethylpyrocarbonate-treated water. The purity and concentration of the total RNA was determined using a NanoDrop1000 (Thermo Scientific, Wilmington, DE, USA), and RNA integrity was verified via electrophoresis (1% agarose gel). The mRNA was reverse-transcribed using a PrimeScript reagent kit (TaKaRa Biotechnology Co., Ltd., China) following the manufacturerŌĆÖs instructions. The reverse transcription products (cDNA) were stored at ŌłÆ20┬░C before being used as templates for quantitative real time-polymerase chain reaction (qRT-PCR). The qRT-PCR was carried out in triplicate in an ABI 7300 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) with a SYBR Premix Ex Taq Kit (TaKaRa Biotechnology Co., Ltd., China). The gene-specific primers are listed in Table 3. The oligonucleotide primers used for the qRT-PCR were designed to span the genomic introns and were synthesized commercially by Invitrogen Biotech Co., Ltd. (Shanghai, China). Thermal cycler conditions consisted of incubation at 95┬░C for 30 s followed by 40 cycles of 95┬░C for 5 s, and then 60┬░C for 31 s. The relative level of target gene expression, as determined by ABI software (Applied Biosystems, USA), was calculated using the comparative 2ŌłÆ╬ö╬öCT method for relative quantification [20]. The mRNA abundance of each target gene was normalized to ╬▓-actin as an internal control (housekeeping gene).
Statistical analysis
All data were analyzed as a mixed model with intrauterine growth type (T) of the pig (NBW or IUGR), content of choline (D) in diets (normal or high), and T├ŚD as fixed effects and the pigs as random effects using the MIXED procedure. If the significance level of the T├ŚD interaction was less than 0.05, a one-way analysis of variance was conducted among the different groups. Pig was considered the experimental unit for all parameters (n = 4) and all of the statistical analyses were performed using SPSS 17.0. The statistical significance was set at p<0.05, and 0.05<p<0.10 was considered a tendency. The data are presented as least squares means with standard error of the mean.
RESULTS
Growth performance
The effects of intrauterine growth type and choline supplementation on the growth performance of pigs is presented in Table 4. The ADG was lower in the IUGR pigs during weaning (d 0 to 21) (p<0.05) compared with the NBW pigs. Although there were not significant differences in ADG after weaning between the NBW and IUGR pigs, IUGR pigs still maintained a lower BW from d 21 to 120. However, we observed that the IUGR pigs showed the similar BW with the NBW pigs (p = 0.07) at d 200. This might be mostly due to the decrease of ADG from d 120 to 200 in NC group compared with the others, although not significant. There were no effects of dietary choline supplementation on BW or ADG, and no T├ŚD interaction was observed.
Serum metabolites
The effects of intrauterine growth type and choline supplementation on the FFA, triglyceride and cholesterol levels of serum in the 200-day-old pigs are shown in Table 5. Increased (p<0.05) FFA and triglyceride levels were observed in the IUGR pigs compared with the NBW pigs. Choline supplementation decreased (p<0.05) the levels of FFAs and triglycerides in the serum of the pigs. There was also a T├ŚD interaction (p<0.01) for the cholesterol level, which was significantly lower (p<0.05) in the NH group than in the other three groups. Although no significant differences in HDL-C and LDL-C were observed among treatments, the ratio of HDL-C to cholesterol was decreased (p<0.05) by IUGR and increased (p<0.05) by a high choline diet. An interaction (p<0.05) between pig and diet was observed in the ratio of HDL-C to cholesterol, which was increased (p<0.05) with choline supplementation in NBW pigs, but not significant in IUGR pigs.
Enzyme activities in the serum and the liver
The effects of choline supplementation and a T├ŚD interaction on LPL activity were both significant (p<0.05) (Table 6). Pigs with IUGR offered the high-choline diet had decreaed LPL activity compared with IUGR offered control diet (p<0.05), but there was no effect of choline supplementation on LPL activity in NBW pigs. The total activity of LPL and HL tended to be reduced (p = 0.09) in the choline-supplemented treatments. The activities of MDH and G6PDH were both increased (p<0.05) in the livers of the IUGR pigs, and choline supplementation decreased (p<0.05) MDH activity (Table 7).
Enzyme and transcriptional factor mRNA expression
The relative mRNA expression of genes related to lipogenesis and lipolysis, as well as transcriptional factors in liver, are shown in Table 8. Compared with the NBW pigs, IUGR pigs had decreased expression of adipose triacylglycerol lipase (ATGL) (p<0.01) and increased mRNA expression of fatty acid synthase (FAS) (p<0.01) and phosphatidylethanolamine N-methyltransferase (PEMT) (p<0.05) in the liver. The FAS, acetyl-CoA carboxylase ╬▒ (ACC╬▒), hormone sensitive lipase (HSL), and PEMT mRNA were expressed at a reduced rate (p<0.05) with choline supplementation. There were T├ŚD interactions (p<0.05) for FAS, ATGL, carnitine palmitoyl transferase 1A (CPT1A), and PEMT expressions. For FAS and PEMT, the results showed that choline supplementation decreased FAS and PEMT mRNA expressions more dramatically (p<0.05) in IUGR pigs than NBW pigs. The T├ŚD interaction for CPT1A mRNA expression indicated that choline supplementation increased CPT1A mRNA expression in IUGR pigs (p<0.05), but did not affect the expression in NBW pigs. The expression of ATGL mRNA was increased (p<0.05) by choline in NBW pigs, but not in IUGR pigs.
DISCUSSION
Choline, as an essential vitamin additive in animal feed mixtures, plays a critical role in cell membrane integrity, lipid metabolism and methylation [21]. The goal of this study was to assess the effects of long-term supplementation of choline on circulating lipid parameters and hepatic lipid metabolism in IUGR pigs. We showed that IUGR pigs demonstrated catch-up growth at d 200 accompanied by abnormal lipid metabolism, and that choline supplementation improved lipid metabolism by decreasing circulating levels of FFAs and triglycerides, as well as LPL activity. Significantly, choline also improved hepatic lipid metabolism by decreasing nicotinamide adenine dinucleotide phosphate (NADPH) generation, down-regulating the expression of lipogenic enzymes and improving fatty acid oxidation, as well as enhancing the export of triacylglycerol from the liver. To our knowledge, this study is the first to show that a diet rich in choline has a role in regulating lipid metabolism in IUGR pigs. These findings provide novel insights into the role of choline in the lipid metabolism of IUGR offspring and provides specific evidence that choline plays different roles in IUGR and NBW pigs.
In the present study, the BW of IUGR pigs remained lower than NBW pigs prior to d 200, but a similar final BW (d 200) was observed for NBW and IUGR pigs. The reduced BW and growth rate in IUGR offspring, particularly in the early stages of life, has been well documented [22]. However, in the case of postnatal exposure to abundant nutrition, IUGR offspring have an opportunity for catch-up growth. A previous study also reported that pigs with low birth weight had a greater weight gain than their heavy littermates at d 105 when cross-fostered to minimize sucking competition and given free access to diets after weaning [23]. In the current study, all pigs were fed ad libitum, and the ADG data showed that catch-up growth began as early as weaning in the IUGR pigs. In addition, as we have found no reports on the effect of high choline on growth performance in pigs, the differences in ADG between NC and NH treatments from d 120 to 200 (although not significant) should be confirmed in the further studies with more experimental animals.
Although it seems beneficial to productive performance, catch-up growth in both IUGR animals and humans is considered an important component of MetS programming [24]. It is well accepted that MetS is generally characterized by blood lipid disorders including high FFA, triglyceride, low HDL-C and high LDL-C levels [25]. Cholesterol homeostasis is an important component of lipid metabolism. HDL-C particles promote vascular health by extracting cholesterol from the tissues (including atherosclerotic plaques) and delivering it back to the liver. Conversely, LDL-C is the classic antagonist of the circulatory system due to its propensity to bind to connective tissue in the intimal sub-layer of the arteries. Thus, increased total cholesterol (particularly LDL-C) and a reduced level of HDL-C are risk factors in the pathogenesis of coronary heart disease and metabolic diseases. In our study, we found that circulating FFA, triglyceride and cholesterol levels were increased and the HDL-C/total cholesterol ratio was decreased in IUGR pigs compared with control pigs, which might indicate a programmed disorder of lipid metabolism in IUGR pigs at d 200.
Choline serves as a structural constituent of membrane phospholipids (PC, choline plasmalogen, and sphingomyelin) and also plays an important role in the other biological actions (cholinergic neurotransmission, platelet activating-factor formation, hepatic secretion of VLDL, and methyl transport). In humans, the beneficial effects of the Mediterranean diet in the prevention of cardiovascular disease were recently associated with a higher intake of choline [26] and subjects with a higher choline intake had a higher HDL-C level [27]. In the present study, we have also shown that high supplementation of choline has a role in increasing the circulating HDL-C/total cholesterol ratio and decreasing the total cholesterol level in the plasma of NBW pigs, but not in IUGR pigs.
LPL is a critical enzyme involved in the hydrolysis of triglycerides carried by chylomicron and VLDL [28], a process which generates FFA for energy metabolism and storage. We observed that choline supplementation decreased circulating LPL activity in IUGR pigs, which might explain the reduced levels of circulating FFAs and triglycerides. The decreased FFA and triglyceride levels in serum suggest an improved lipid metabolism status in the liver, as increased circulating FFAs and triglycerides are important component of fatty liver, liver insulin resistance and type II diabetes [29].
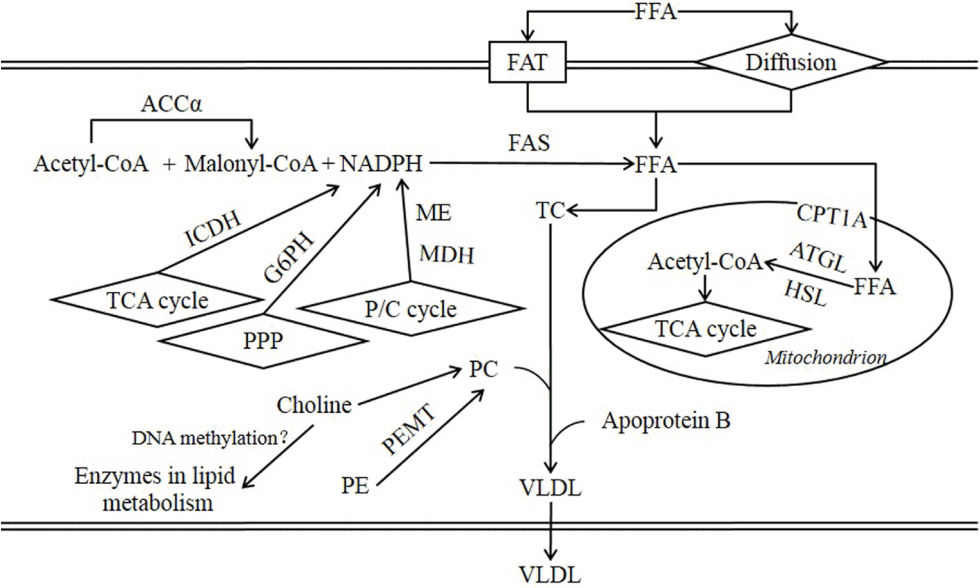
With the exception of FFA and triglyceride uptake, the hepatic lipid metabolism is primarily involved in the de novo synthesis and oxidation of fatty acids, as well as lipoprotein synthesis and export (Figure 1). In brief, de novo lipogenesis in the liver begins with the carboxylation of acetyl-CoA to malonyl-CoA through the actions of ACC╬▒. FAS then converts malonyl-CoA to its primary product, palmitic acid. The NADPH-generating enzymes, such as ME, MDH, ICDH, and G6PDH, provide the primary sources for lipogenesis. CPT1A controls the transport of long-chain acyl-CoA into the mitochondria, which is the rate-limiting step of mitochondrial fatty acid ╬▓-oxidation in the liver. Recent studies have found that the first step in the oxidation of fatty acids is the mobilization of triacylglycerol, which is catalyzed by ATGL and HSL [30]. Although normally presenting at very low levels in the liver, ATGL and HSL both appear to channel the liberated fatty acids towards oxidation [31]. In the present study, we observed that the mRNA expression of FAS, and the activities of MDH and G6PDH in the liver were increased, and ATGL expression was decreased by IUGR. With a normal dietary choline content, the expression of CPT1A was also decreased in IUGR pigs compared with NBW pigs in this study, which is consistent with the existing literature on rats [32]. Because of these findings, we speculate that for IUGR pigs, there was abundant NADPH and more efficient enzymes for the de novo lipogenesis of fatty acids, but that the oxidation of fatty acids might be decreased, causing the accumulation of lipid droplets in the hepatocytes. The current study also shows that choline supplementation down-regulated hepatic MDH activity and mRNA expression of FAS and ACC╬▒, but its effect of up-regulating hepatic CPT1A mRNA expression was only observed in IUGR pigs and not in NBW pigs. The increased expression of CPT1A is consistent with previous studies [33], which reported the fatty acid oxidation-promoting effect of choline in the liver of both guinea pigs and humans. These alterations might reduce triglyceride accumulation in the liver and improve liver function, as indicated by decreased serum AST and ALT concentrations in the present study in response to choline supplementation in IUGR pigs.
As the circulating FFA level and hepatic expression of FAS mRNA were increased by IUGR, more PC was needed to package and export triacylglycerol in VLDL to avoid the accumulation of lipid in the liver. PC is synthesized in the liver through two pathways [34]: i) the cytidine diphosphate (CDP) -choline pathway with dietary choline as the substrate; and ii) the methylation of phosphatidylethanolamine catalyzed by PEMT. When the dietary choline was adequate to synthesize abundant PC, PEMT expression and activity are compensatorily down-regulated [35], which was confirmed in IUGR pigs in the current study. The choline supplementation down-regulated the gene expression of PEMT in the liver of IUGR pigs, suggesting stimulating of the CDP-choline pathway to synthesize PC in the IUGR pigs. However, the expression of PEMT mRNA was not altered by choline administration to NBW pigs. These inconsistencies might be attributed to a difference in choline metabolism between IUGR and NBW pigs, as a different choline metabolism status has been observed in people with a different metabolic status [36]. Because IUGR caused a decrease in the circulating level of choline in pigs after weaning [10], we speculate that the high concentration of choline in the supplemented diet helped IUGR pigs to maintain normal circulating levels of choline and adequate PC synthesis.
Sterol regulatory element-binding proteins 1c (SREBP-1c) is one of the most important transcription factors in the liver for regulating de novo fatty acid synthesis through transcriptionally-mediated activation by insulin. The mitochondrial, peroxisomal, and microsomal fatty acid oxidation systems are regulated by peroxisome proliferator-activated receptor alpha in the liver. However, in the present study, transcription factors were not significantly altered by IUGR or choline supplementation. Although the increased expression of fat-synthesizing enzymes might be induced by serum triglycerides [37], the increased expression of FAS in this study was independent of the typical inducers (i.e., circulating insulin and glucose, data not presented) and the mediator SREBP-1c, suggesting that liver lipid metabolism in IUGR offspring may result from metabolic programming [38]. Although it has been suggested that dietary choline intake is an important modifier of the epigenome [39], the effects of long-term supplementation of high levels of choline on metabolic programming and DNA methylation remain to be explored.
In summary, this study demonstrated that finishing pigs with IUGR had abnormal circulating lipid parameters compared with NBW pigs, and that diets with high levels of choline might have a role in regulating hepatic lipid metabolism through decreasing activity of NADPH-generating enzymes and altering expressions of lipid metabolism-related genes. Our data provide evidence that treating IUGR offspring after birth with dietary supplementation that includes high levels of choline improves lipid metabolism in the liver. Further research is needed to determine the levels of choline and its metabolites in the liver and in the circulation of IUGR pigs in large groups.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print







