




INTRODUCTION
Nutritional value of protein in feedstuff not only depends on its amino acid content and composition proportion, but also on its molecular structure characteristics (Peng et al., 2014). Recent studies have highlighted the importance of determining the relationship between molecular spectral band characteristics of proteins and their intestinal absorption so as to clarify the mechanism of nitrogen supplementation (Qin et al., 2014). The traditional method of determining the nutritional value of feeds involves destroying the three-dimensional structure of constituent proteins by acid or alkali treatment or by mechanical polishing, and then using so-called wet chemical methods to determine chemical composition. However, this approach provides no information at a molecular level on the nutritional value of feeds or on their capacity for digestion by livestock. Fourier transform infrared (FTIR) technique is a powerful tool for analyzing the structure of feed proteins (Xin et al., 2013a), which allows investigators to predict how they are processed and absorbed within the animalsŌĆÖ digestive tract. However, there is certainly difficulty to use FTIR spectrum for quantifying the tertiary structure having higher portion of side chains of the conventional feedstuffs protein.
Recent studies have shown a close correlation between the molecular structure characteristics of feed proteins and their nutritional value (Yu and Nuez-Ort├Łn, 2010; Samadi et al., 2013). Specifically, ╬▒-helices and ╬▓-sheets in the protein secondary structures can affect digestibility of feed proteins (Dyson and Wright, 1993). Certain features (amide I and II bands) of protein molecular structures of corn distillerŌĆÖs dried grains with solubles (corn DDGS) from various sources were correlated with intestinal protein digestibility, which can be used to predict the absorption of essential nutrients by the small intestine (Chen et al., 2014). Another study found that spectral band intensity characteristics of proteins affect protein quality, nutrient utilization, and digestion in dairy cows (Doiron et al., 2009). Therefore, establishing a relationship between the molecular structure characteristics of conventional feed proteins and their nutritional value will strengthen the current knowledge on protein nutritional value and broaden the scope of molecular spectroscopy as a bioanalytical tool for rapidly assessing the nutritive value of feed proteins.
To our knowledge, the relationship between molecular structure characteristics of conventional feed proteins and protein solubility (PS) and in vitro digestibility in monogastric animals has not been previously investigated. The objectives of this study were i) to assess protein molecular structure characteristics of conventional feed proteins by FTIR; ii) to examine the PS and in vitro digestibility of these proteins; and iii) to quantify the relationship between protein molecular structure characteristics and PS as well as in vitro digestibility.
MATERIALS AND METHODS
Sample preparation and treatment
Five conventional feed ingredients (soybean meal, fish meal, corn DDGS, corn gluten meal, and feather meal) were provided by Changchun Wellhope Animal Husbandry Co. (Changchun, China). All feed samples used in this experiment were collected from raw commodities and stored according to commodity storage standard, one month of storage period. One half of each sample was ground in the laboratory mill through a 1 mm screen for laboratory analysis and sub-samples (n = 3 per type of feed) was collected and stored at 4┬░C. The other half was stored as a backup at ŌłÆ20┬░C. Chemical profiles of the five feeds are presented in Table 1.
Determination of protein solubility
PS was determined according to the method as described by Dale (1987). Feedstuff sample with 1.5 g was placed in a 100 mL conical flask, and then added 75 mL of 0.2% (w/v) potassium hydroxide solution; the mixture was stirred on a magnetic stirrer for 20 min and then transferred to a 50 mL tube and centrifuged at 2,700 rpm for 10 min. 15 mL volume of supernatant was measured by the chemical method (AOAC, 1990), yielding an amount equivalent to 0.3 g of original sample (15/75 = x/1.5 g, where x = 0.3 g). PS (%) was calculated as crude protein (CP) content in 15 mL supernatant/CP content of the original sample├Ś100%.
Determination of protein in vitro digestibility
Protein in vitro digestibility was evaluated according to a method as previously described by Boisen and Fernandez (1995). Feedstuff sample with 1.0 g (accurate to 0.001 g) was placed in a 100 mL conical flask, and then added 10 mL of 0.01 M hydrochloric acid (pH 2.0) and pepsin solution containing 1.0 mg porcine pepsin (product no. P7-000; Sigma, St. Louis, MO, USA). To prevent bacterial growth, 0.5 mL of chloramphenicol solution consisting of 0.5 g chloramphenicol (product no. 0230; Amresco, Solon, OH, USA) in 100 mL ethanol was added to the mixture, followed by incubation for 4 h at 37┬░C. After neutralization with 0.2 M sodium hydroxide, 10 mL of 0.2 M phosphate buffer (pH 6.8) was added. The pH was adjusted to 6.8 with 1 M HCl or 1 M NaOH, and the solution was mixed with 1 mL of a freshly prepared pancreatin solution containing 50 mg porcine pancreatin (product no.P7-545-100G; Sigma, USA). The flask was closed with a rubber stopper and incubated with continuous magnetic stirring at 39┬░C for 4, 8, 12, 16, 20, 24, and 28 h, respectively. After adding 5 mL of 20% sulfosalicylic acid, the sample was centrifuged at 15,000 rpm for 15 min; the supernatant was discarded and the precipitate was heated at 80┬░C for 24 h. CP digestibility (in %) was calculated as CP content of the original sample ŌłÆCP content of precipitate/CP content of the original sample├Ś100%.
Fourier transform infrared spectroscopy
FTIR spectroscopy was performed according to the method as described by Long et al (2015). Briefly, 2 mg of sample was mixed with 200 mg KBr in a vacuum dryer for 24 h; the mixture was uniformly ground and pressed into a tablet (tablet thickness = 0.25 mm; translucent shape) using an infrared (IR) tablet press (FW-4; Thermo Fisher Scientific, Waltham, MA, USA). FTIR spectra (FTIR-8400s; Shimadzu, Kyoto, Japan) were acquired in the mid-IR range (4,000 to 400 cm ŌłÆ1) at a resolution of 4 cm ŌłÆ1 with 64 co-added scans. Each sample was run six times. As a control, a KBr pellet without protein was recorded under identical conditions. IR spectra were processed using OMNIC 8.0 software (Nicolet Analytical Instruments, Madison, WI, USA), and after baseline correction of the original map the sample and control spectra were compared. Amide I band narrowing was achieved with a full height of 38.2 and a resolution enhancement factor of 4.4. Fourier self-deconvolution using Origin 7.5 software (Origin Lab, Northampton, MA, USA) was used for Gaussian curve fitting in the region of the amide I band to separate overlapping bands. The secondary structure content of samples was detected from IR second-derivative amide I spectra by manually computing the relative peak areas under the bands assigned to a particular substructure.
Statistical analysis
Data were evaluated by analysis of variance with SPSS v.19.0.1 software (SPSS Inc., Chicago, IL, USA). The model used for protein molecular spectral band characteristics, PS and in vitro digestibility analysis was Yij = ╬╝i+eij, where Yij is an observation on the dependent variable ij (i = 1, 2,. . .,5; j = 1, 2, 3), ╬╝i is the overall mean (i = 1, 2,. . .,5), and eij is the random error associated with the observation ij. Multiple comparisons of group means were carried out using DuncanŌĆÖs method. Results are expressed as mean┬▒standard error, with ╬▒ = 0.05 indicating a statistically significant difference. Correlation analysis was performed using PearsonŌĆÖs correlation coefficient with 95% confidence limits (p<0.05).
RESULTS
Protein spectral band intensities
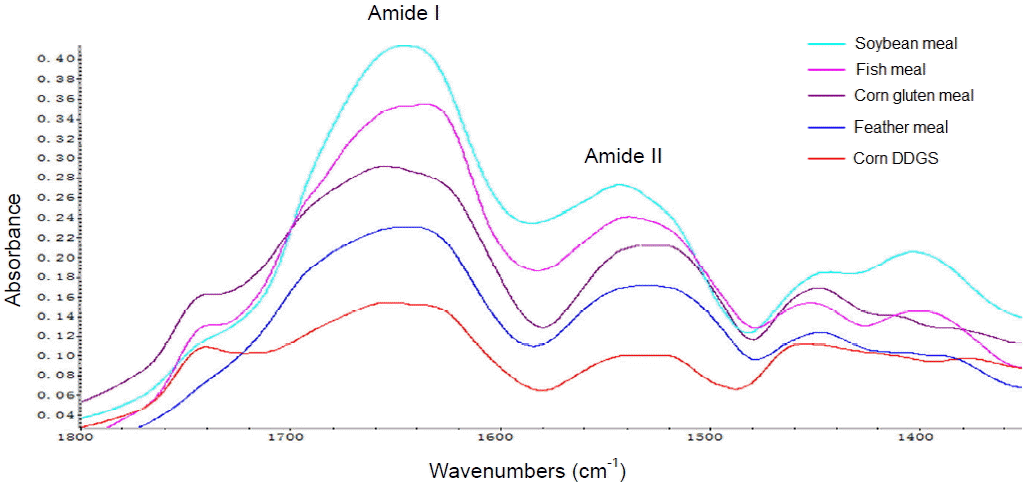
The protein IR spectrum showed two salient features, i.e., the amide I (1,700 to 1,600 cmŌłÆ1) and amide II (1,600 to 1,500 cmŌłÆ1) bands (Figure 1). Results of the protein spectral band intensities analyses are presented in Table 2. Significant differences were observed among feeds in terms of amide II heights and areas and amide I-to-II height and area ratios (p<0.001). Soybean meal had the highest values for amide I and II heights, amide I and II areas, and amide I-to-II height and area ratios, whereas corn DDGS had the lowest values for all of these features.
Protein secondary structures and solubility
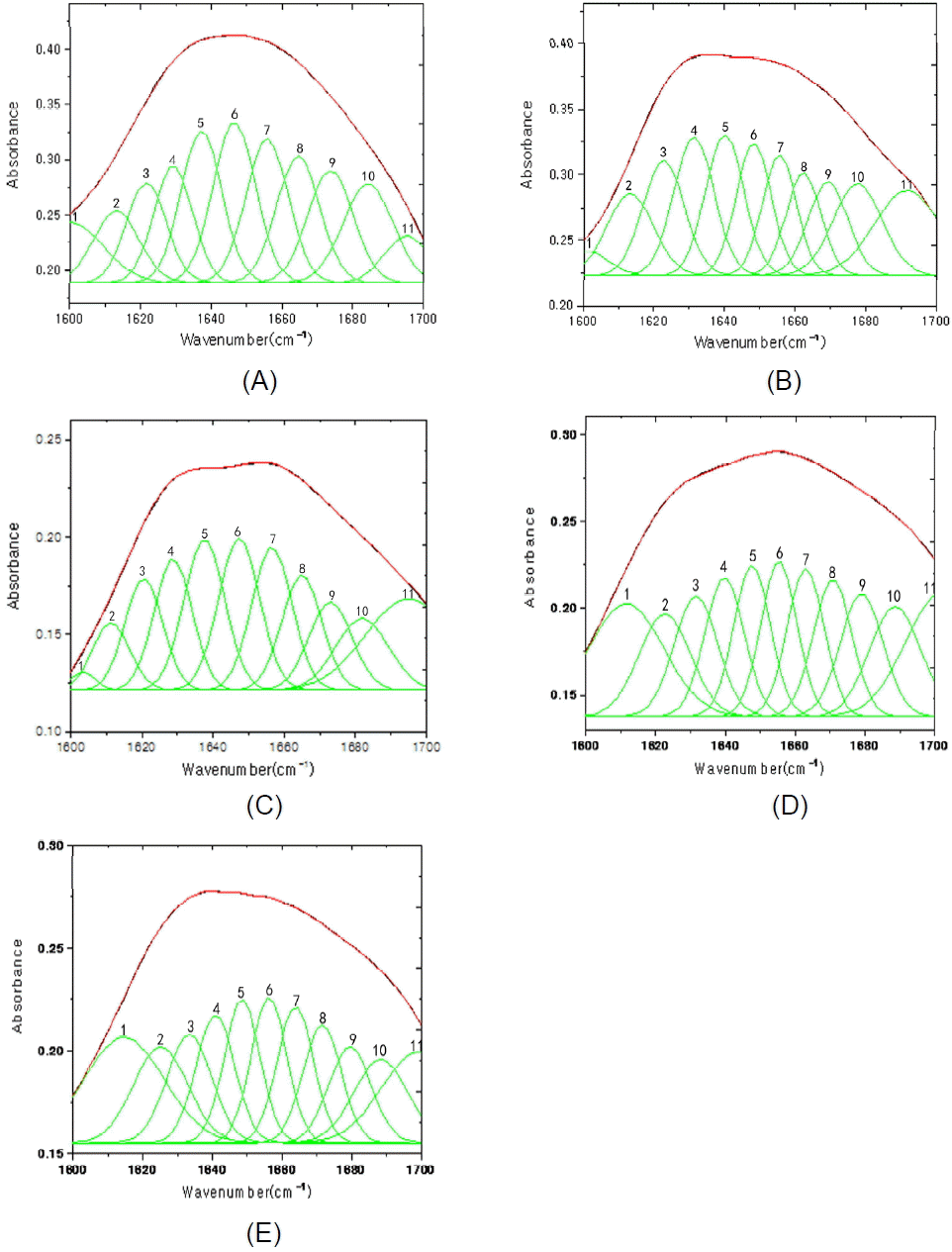
The Gaussian curve fitting analysis of the amide I band peak was carried out based on the preliminary study of Meng-Xia and Yuan (2002), showed characteristic peaks at 1,611 to 1,639 cmŌłÆ1 that were attributed to ╬▓-sheets. Other peaks were observed that corresponded to the following structures: random coils, 1,640 to 1,649 cmŌłÆ1; ╬▒-helices, 1,650 to 1,658 cmŌłÆ1; and ╬▓-turns, 1,660 to 1,700 cmŌłÆ1 (Figure 2). Results of the protein secondary structures and solubility analyses are shown in Table 3. Soybean meal had the highest percentage of ╬▒-helices and random coil structures and ╬▒-helix-to-╬▓-sheet ratios; feather meal had the highest percentage of ╬▓-sheet structures; and corn DDGS had the highest percentage of ╬▓-turn structures, while corn gluten meal had the lowest percentage of ╬▓-turn and random coil structures. The rank order of PS was soybean meal>fish meal>corn DDGS>corn gluten meal>feather meal (p<0.05).
Protein in vitro digestibility
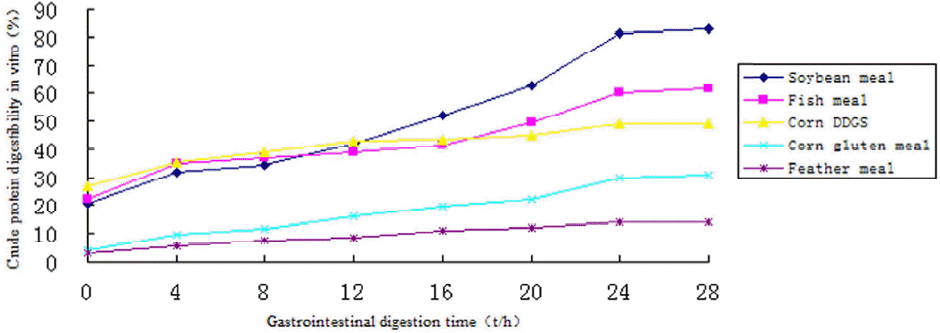
Evaluation of protein in vitro digestibility revealed significant differences among five conventional feed ingredients at each time point (p<0.05; Figure 3). The in vitro digestibility of proteins by pepsin-trysin increased over time. Soybean and feather meal had the highest and lowest in vitro digestibility values, respectively, at 28 h. The rank order of protein in vitro digestibility at this time point was soybean meal>fish meal>corn DDGS>corn gluten meal>feather meal (p<0.05). The lower in vitro digestibility of corn DDGS proteins at 28 h as compared to that at 24 h may be explained by the inhibition of enzymatic reactions due to accumulation of digestion products.
Correlation between protein spectral band intensities and protein in vitro digestibility and solubility
Coefficients of correlation between protein spectral band intensities (height and area) and protein in vitro digestibility at 28 h as well as solubility are shown in Table 4. Amide I height was positively correlated with PS (r = 0.958, p<0.001) and in vitro digestibility (r = 0.848, p< 0.001), as were amide II height (r = 0.707, p = 0.003 and r = 0.518, p = 0.048, respectively), the height ratio of amide I to II (r = 0.857, p<0.001 and r = 0.894, p<0.001, respectively), amide I area (r = 0.959, p<0.001 and r = 0.959, p<0.001, respectively), amide II area (r = 0.709, p = 0.003 and r = 0.519, p = 0.047, respectively), and area ratio of amide I to II (r = 0.854, p<0.001 and r = 0.893, p<0.001, respectively).
Correlation between protein secondary structures and protein in vitro digestibility and solubility
Coefficients of correlation between protein secondary structures and protein in vitro digestibility at 28 h as well as solubility are shown in Table 5. The percentage of ╬▒-helix structures was positively correlated with PS (r = 0.903, p<0.001) and in vitro digestibility (r = 0.916, p<0.001), as were the percentage of random coil structures (r = 0.694, p = 0.004 and r = 0.789, p<0.001, respectively) and ╬▒-helix-to-╬▓-sheet ratios (r = 0.955, p<0.001 and r = 0.966, p<0.001, respectively). However, the percentage of ╬▓-sheet structures was negatively correlated with both protein in vitro digestibility (r = ŌłÆ0.826, p<0.001) and solubility (r = ŌłÆ0.740, p = 0.002).
DISCUSSION
There is an increasing demand to improve the nutritional value of feed proteins for monogastric animals by optimizing feed protein sources and increasing the efficiency of protein utilization, which requires a fast and accurate estimation of the intestinal digestibility of feed proteins. Earlier findings suggest that the pepsin-pancreatin two-step enzymatic method is potentially a useful for evaluating in vivo digestibility (Boisen et al., 1995). However, previous studies have been unable to explain the variability in the digestion of different feed proteins, which are likely due to differences in protein structure.
The present study investigated the molecular spectral band characteristics of proteins in various types of feedstuff in relation to their digestibility and solubility. PS and protein spectral band intensities (height and area) in the feedstuff examined in this study varied widely. Protein secondary structures such as ╬▒-helices, ╬▓-sheets, and random coils as well as ╬▒-helix-to-╬▓-sheet ratios in soybean meal and corn DDGS, and the relative quantitative amounts of ╬▒-helices and ╬▓-sheets in feather meal were inconsistent with earlier findings (Yu et al., 2004; Jiao et al., 2012). Even the reason for these differences is unclear, they may be related to variations inherent in feed sources, processing technology, or storage conditions (Peng et al., 2014).
We found that in vitro digestibility of soybean meal protein was lower than previously reported values (Boisen et al., 1995), which may have been due to the presence of anti-nutritional factors (Tan-Wilson et al., 1985) that can inhibit enzymatic reactions. On the other hand, fish meal protein in vitro digestibility was higher than that reported in another study (Huang et al., 2000) whereas feather meal protein in vitro digestibility was similar to results obtained by other investigators (Yu et al., 2004). This may be due to the presence of keratinŌĆöa scleroprotein that makes up 85% to 90% of feather mealŌĆöwhich has highly stable S-S and SH linkages that are not readily broken down by animals without processing (Papadopoulos et al., 1986). One possible reason for which corn DDGS and gluten meal had lower protein in vivo digestibility and PS is their low solubility in water due to a high content of alcohol-soluble proteins containing hydrophobic amino acids (Liu et al., 2013); alternatively, the S-S or O-H bonds in their structures may promote the formation of ╬▒-helices (Zhang and Yu, 2012), which are structurally stable and therefore not easily digested.
The amide I band mainly arises from stretching vibrations of the amide C=O group, while the amide II band is attributable to N-H bending (60%) and C-N stretching (40%) vibrations; these have also been used to determine protein structure or conformation (Yan et al., 2014), and the protein two primary spectral band intensities in terms of peak height and peak area indicate quantitative differences in protein functional groups (Peng et al., 2014). Corn DDGS and feather meal degraded slowly as compared to the other types of feed due in part to their low basic amino acid content, which is indirectly reflected by amide height and area. Trypsin is the main protease in the intestine and targets the peptide bond between lysine and arginine; therefore, the lack of basic amino acids would diminish the probability of contact between the CP in feed and digestive enzymes in the animal, thereby decreasing the amount and variety of oligopeptides and free amino acids that are released (Adler-Nissen, 1979).
The vibrational frequency of the amide I band is particularly sensitive to and can be used to predict protein secondary structure (Yu et al., 2004). Our findings confirmed that the percentage of ╬▓-sheet structures is closely related to the nutritional value of feed, with higher percentage being associated with lower PS and CP digestibility in the small intestine, since ╬▓-sheets have a large number of hydrogen bonds that can hinder protease activity. We also confirmed that the nutritional value of proteins feed differed according to ╬▒-helix-to-╬▓-sheet ratio; previous studies have also reported that this ratio is a good predictor of nutritional value or digestibility of proteins feed (Theodoridou and Yu, 2013; Xin et al., 2013b). We found that the percentage of random coil structures was positively correlated with PS and digestibility, possibly because this structure is strong and flexible and has fewer hydrogen bonds that can impede enzyme access, which allows proteins to be readily degraded.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







