Influence of Palm Kernel Meal Inclusion and Exogenous Enzyme Supplementation on Growth Performance, Energy Utilization, and Nutrient Digestibility in Young Broilers
Article information
Abstract
The objective of the present study was to investigate the influence of palm kernel meal (PKM) inclusion and exogenous enzyme supplementation on growth performance, nitrogen-corrected apparent metabolizable energy (AMEn), coefficient of apparent ileal digestibility (CAID) and total tract retention of nutrients in young broilers fed corn-based diets. Four inclusion levels of PKM (no PKM [PKM0], 8% [PKM8], 16% [PKM16], and 24% [PKM24]) and two enzyme additions were evaluated in a 4×2 factorial arrangement of treatments. A total of 384, one-d-old male broilers (Ross 308) were individually weighed and allocated to 48 cages (eight broilers/cage), and cages were randomly assigned to eight dietary treatments. Results indicated that the inclusion of 8% and 16% PKM increased (p<0.05) the weight gain compared to the PKM0 diet. Birds fed the PKM8 diets had the highest (p<0.05) feed intake. Weight gain and feed intake were severely reduced (p<0.05) by feeding the PKM24 diet. Enzyme supplementation increased weight gain (p<0.05), independent of PKM inclusion level. In PKM0 and PKM8 diets, enzyme addition significantly (p<0.05) lowered feed conversion ratio (FCR); whereas enzyme addition had no effect on FCR of birds fed PKM16 and PKM24 diets. In PKM0 and PKM16 diets, enzyme addition significantly (p<0.05) increased CAID of nitrogen and energy but had no effect in the PKM8 and PKM24 diets. Inclusion of PKM into the basal diet, irrespective of inclusion level, enhanced (p<0.05) starch and fat digestibility. Inclusion of PKM at 16% and 24% resulted in similar CAID of neutral detergent fiber (NDF) but higher (p<0.05) than that of the PKM0 and PKM8 diets. Enzyme addition, regardless of the level of PKM inclusion, significantly (p<0.05) increased CAID of NDF. There was a significant (p<0.05) decrease in AMEn with PKM inclusion of 24%. The present data suggest that inclusion of PKM in broiler diets could be optimized if PKM-containing diets are formulated based on digestible amino acid contents and supplemented with exogenous enzymes. If amino acid digestibility and AME of PKM considered in the formulation, it can be included in broiler diets up to 16% with no deleterious effects on growth performance.
INTRODUCTION
High demand for conventional feed ingredients due to competition with humans and other livestock species for feed resources and ever-increasing cost of these ingredients have motivated poultry nutritionists to maximize the use of locally available by-products. Palm kernel meal (PKM), an agro-industrial by-product, is a locally available and relatively inexpensive feedstuff in many tropical countries (Perez et al., 2000). Palm kernel meal is produced by extracting the oil from palm kernels using solvent extraction. Palm kernel oil might also be mechanically expelled and the coproduct is referred to as palm kernel cake (PKC; Sundu et al., 2006). Incorporation of PKM in broiler diets is limited and reported to be associated with impaired performance parameters (Mardhati et al., 2011). The deteriorated growth responses has been attributed to high fiber content, grittiness (Sundu et al., 2005; 2006) and low concentration of indispensable amino acids (AA) (Abdollahi et al., 2015).
Published data on the effect of PKM on growth performance of broiler chickens have been contradictory. Panigrahi and Powell (1991) reported that if supplemented with methionine and lysine, PKM can be included in broiler diets up to 40%. Ezieshi and Olomu (2008) replaced 50% of corn in the broiler diet with PKM (30% and 32.5% PKM in starter and finisher diets, respectively) and reported lower body weight and a deteriorated feed conversion ratio (FCR). Shakila et al. (2012) reported that PKM up to 10%, with or without enzyme, might be used in broiler diets without negatively affecting the performance.
Palm kernel meal contains high level of fiber with non-starch polysaccharides (NSP) and β-mannan as the main fiber components of PKM. Almaguer et al. (2014) reported concentrations of 494 and 779 g/kg of acid detergent fiber and neutral detergent fiber (NDF) in PKM. Knudsen (1997) reported values of 466 and 136 g/kg for NSP and lignin in PKM, respectively. Abdollahi et al. (2015) reported that almost 96% of total NSP in PKM being in the form of insoluble NSP. Beta-mannans, also known as β-galactomannans, are linear polysaccharides with repeating units of β-1-4 mannose and side attachments of α-1-6 glucose and/or galactose and commonly found in many feed ingredients (Knudsen, 1997; Jackson et al., 2004). Hsiao et al. (2006) reported β-mannan concentration of 12.6 and 16.1 g/kg in dehulled and non-dehulled soybean meal samples, respectively. Mannans in PKM are hard, highly-insoluble crystalline polymers of mannose and constitute the storage polysaccharides of the endosperm of palms (Daud and Jarvis, 1992). According to Dusterhoft et al. (1991), 78% of the total NSP in PKM is linear mannan. Jackson (2010) reported that PKM has a β-mannan content of 300 to 350 g/kg. Mannans are not depolymerised in the digestive tract of the birds due to the lack of mannan-degrading enzymes.
Poultry are not able to degrade NSP due to lack of endogenous fiber-degrading enzymes and therefore feeding high fiber ingredients such as PKM might impair growth responses. Fiber in PKM has been found to have a complex structure consisting mannans (glucomannan and galactomannan) as the main component, cellulose, glucoronoxylans and arabinoxylans (Dusterhoft et al., 1991; Knudsen, 1997). Due to its complexity, fiber of PKM may benefit more from a combination of enzymes including β-mannanase, xylanase, cellulose, glucosidase and galactosidase. However, studies investigating the effect of enzyme on PKM-containing broiler diets are limited (Soltan, 2009; Shakila et al., 2012). This study sought to investigate the efficacy of an enzyme cocktail in corn-soybean meal-based broiler diets with graded levels of PKM. The objective of the current study was to examine the effects of four dietary inclusion levels of PKM, each without and with enzyme supplementation, on growth performance, nutrient digestibility and energy utilization of young broilers.
MATERIALS AND METHODS
Enzyme
The enzyme was supplied by AsiaPac (Dongguan) Biotechnology Co. Ltd, Guangdong, China, and was a cocktail of β-mannanase and NSP-degrading enzymes (Pokazyme PK516; including: β-mannanase, xylanase, amylase, protease, cellulase and β-glucanase). The enzyme was in powder form and added at the level recommended by the manufacturer (200 g/t of feed).
Diets
The experimental design was a 4×2 factorial arrangement of treatments, which included four different levels of PKM inclusion (0%, 8%, 16%, and 24%) and two enzyme supplementations (without/with enzyme). Whole corn was obtained from a commercial supplier, and ground in a hammer mill (Bisley’s Farm Machinery, Auckland, New Zealand) to pass through a screen size of 4.0 mm. Broiler starter diets, based on corn, soybean meal and PKM, were formulated to meet the Ross 308 strain recommendations for major nutrients (Ross, 2007) and to be equivalent in respect of energy density, and digestible protein and AA, calcium and available phosphorus concentrations. Palm kernel meal used in the current study had previously been analysed for nutrients, apparent metabolizable energy (AME) and ileal AA digestibility (Abdollahi et al., 2015) and the data were used to formulate the experimental diets. Four diets were formulated to contain no PKM (PKM0), 8% (PKM8), 16% (PKM16), and 24% (PKM24) PKM (Table 1) and used to develop eight dietary treatments without and with the addition of enzyme. The diets contained 0.3% of titanium dioxide (TiO2, Merck KGaA, Darmstadt, Germany) as an indigestible marker for the determination of ileal nutrient digestibility. Diets were mixed in a single-screw paddle mixer (Bonser Engineering Co. Pty. Ltd., Merrylands, Australia). Following mixing, all diets were steam-conditioned at 70°C and pelleted using a pellet mill (Model Orbit 15; Richard Sizer Ltd., Kingston-upon-Hull, UK) capable of manufacturing 180 kg of feed/h and equipped with a die ring with 3-mm holes and 35-mm thickness. Conditioning time of the mash was 30 s and the conditioning temperature was measured at the outlet of the conditioner. The diets were run in sequence with no change in the feeder rate, rotation speed or number of knives. Representative samples of all diets were collected after pelleting for chemical analysis.
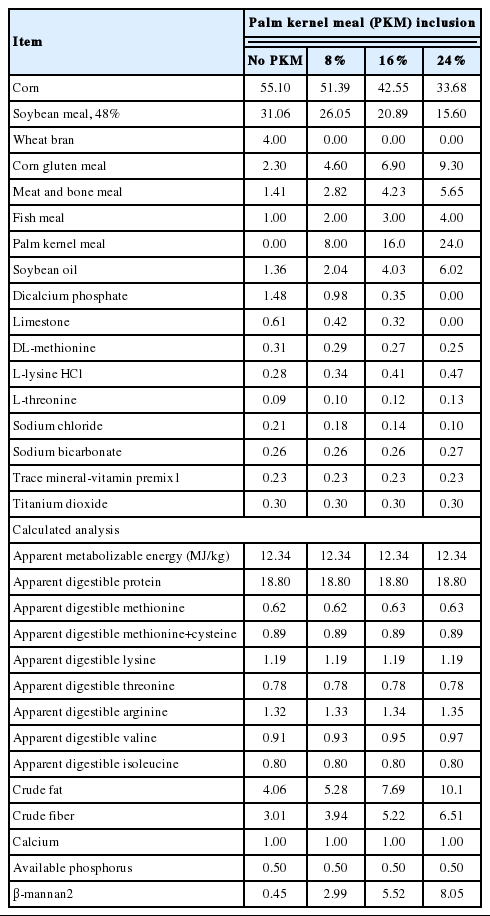
Composition and calculated analysis (%, as fed) of the experimental diets
Birds and housing
Experimental procedures were approved by the Massey University Animal Ethics Committee and complied with the New Zealand Code of Practice for the Care and Use of Animals for Scientific Purposes. One-d-old male broilers (Ross 308), obtained from a commercial hatchery, were individually weighed and allocated to 48 cages (eight birds per cage) in electrically heated battery brooders so that the average bird weight per cage was similar. Six cages were then randomly assigned to each of the eight dietary treatments. The birds were transferred to grower cages on d 12 and fed the same diets until d 21. The space allocation per bird in brooder and grower cages was 530 and 640 cm2, respectively. The battery brooders and grower cages were housed in an environmentally controlled room with 20 h of fluorescent illumination per day. The temperature was maintained at 31°C on d 1, and was gradually reduced to 22°C by 21 d of age. Feed was offered ad libitum and water was freely available.
Pellet durability
Pellet durability was determined in a Holmen Pellet Tester (New Holmen NHP100 Portable Pellet Durability Tester, TekPro Ltd., Willow Park, North Walsham, Norfolk, UK) using the method described by Abdollahi et al. (2010). Clean pellet samples (100 g; four replicates per diet), with no fines, were rapidly circulated in an air stream around a perforated test chamber for 30 s. Fines were removed continuously through the perforations during the test cycle. After the test cycle, the subject pellets were ejected and weighed manually. The pellet durability index (PDI) was calculated as the percentage of the pellets not passing through the perforations at the end of the test.
Performance data
Body weight and feed intake were recorded on a cage basis at weekly intervals. Mortality was recorded daily. Feed conversion ratio values were corrected for the body weight of any bird that died during the course of the experiment.
Apparent metabolizable energy determination and excreta scoring
Feed intake and total excreta output of each cage were quantitatively measured from d 18 to 21 post-hatch. Daily collections from each cage were pooled, mixed in a blender and sub-sampled. Sub-samples were lyophilized (Model 0610, Cuddon Engineering, Blenheim, New Zealand) ground to pass through a 0.5 mm sieve and stored in airtight plastic containers at 4°C pending analysis. The diets and excreta samples were analysed for dry matter (DM), gross energy (GE), nitrogen (N), fat and NDF. On d 21, excreta were also scored for stickiness on a scale of 1 to 5, with 1 representing normally formed excreta and 5 representing watery and very sticky excreta.
Coefficient of ileal apparent digestibility determination
On d 21, six birds per cage were euthanized by intravenous injection (1 mL per 2 kg live weight) of sodium pentobarbitone solution (Provet NZ Pty Ltd., Auckland, New Zealand) and ileal digesta were collected as described by Ravindran et al. (2005). The ileum was defined as that portion of the small intestine extending from the Meckel’s diverticulum to a point ~ 40 mm proximal to the ileo-caecal junction. The ileum was then divided into two halves and the digesta was collected from the lower half towards the ileo-caecal junction. Digesta from birds within a cage were pooled, lyophilized (Model 0610, Cuddon Engineering, Blenheim, New Zealand), ground to pass through a 0.5 mm sieve and stored at 4°C until laboratory analysis. The diets and digesta samples were analysed for GE, titanium dioxide (Ti), N, starch, fat, and NDF.
Chemical analysis
Dry matter was determined using standard procedures (method 930.15; AOAC, 2005). Gross energy was determined by adiabatic bomb calorimetry (Gallenkamp Autobomb, London, UK) standardized with benzoic acid. Samples were assayed for Ti on a UV spectrophotometer following the method of Short et al. (1996). The NDF was determined using standard procedures (AOAC, 2005).
Nitrogen was determined by combustion (method 968.06; AOAC, 2005) using a CNS-200 carbon, N and sulphur auto analyser (LECO Corporation, St. Joseph, MI, USA). Starch was determined using the Megazyme Total Starch Assay kit (Megazyme International Ireland Ltd., Wicklow, Ireland) based on thermostable α-amylase and amyloglucosidase. Fat was determined using Soxhlet extraction procedure (method 991.36; AOAC, 2005).
Calculations
All data were expressed on a DM basis, and the AME was calculated using the following formula:
Nitrogen-corrected AME (AMEn) was determined by correction for zero N retention by simple multiplication with 36.54 kJ per gram N retained in the body as described by Hill and Anderson (1958).
Coefficient of apparent ileal digestibility (CAID) of nutrients was calculated using the following formula:
Where (Diet component/Ti)diet = ratio of diet component to Ti in the diet, and
Total tract retention (TTR) of nutrients, as a percentage of intake, was determined as follows:
Statistical analysis
The data were analysed by two-way analysis of variance to determine the main effects (PKM inclusion and enzyme addition) and their interaction using the General Linear Models procedure of SAS (2004). Cage means served as the experimental unit for all data. Differences were considered to be significant at p<0.05 and significant differences between means were separated by the Least Significant Difference test.
RESULTS
Pellet durability index
Increasing the dietary PKM inclusion resulted in lower PDI compared to the diet with no PKM. The PDI values of 85.3, 56.2, 34.4 and 15.4, respectively, were obtained with PKM inclusion of 0%, 8%, 16%, and 24%.
Performance and, excreta score and dry matter
Mortality during the performance experiment was negligible. Only six out of the 384 birds died and the deaths were not related to any specific treatment.
The performance data of broiler starters from d 1 to 21 post-hatch is shown in Table 2. Weight gain of the birds was significantly (p<0.001) influenced by PKM inclusion and enzyme addition. Inclusion of 8.0% and 16% PKM increased (p<0.05) the weight gain compared to the PKM0 diet. Weight gain was severely reduced by feeding the PKM24 diets. Enzyme supplementation increased weight gain (p<0.05), irrespective of PKM inclusion level.
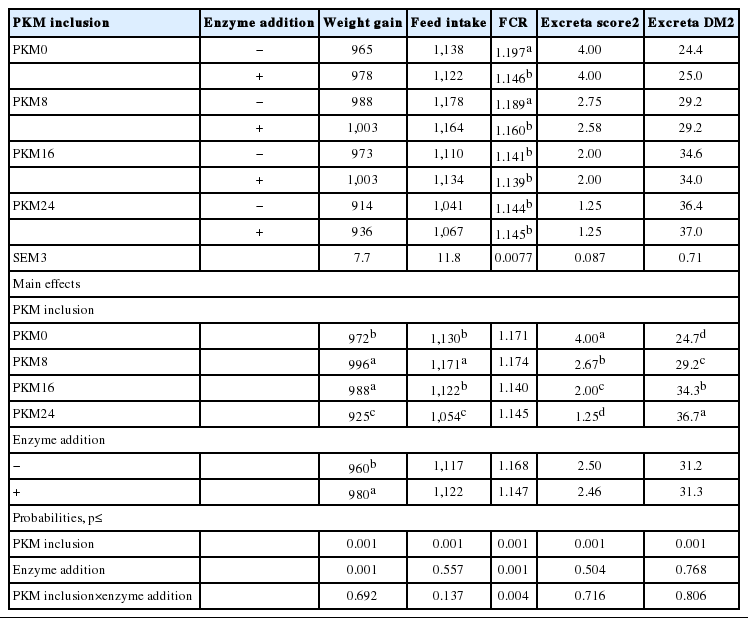
There was a significant (p<0.001) effect of PKM inclusion on feed intake with birds fed the PKM8 diets had the highest feed intake (p<0.05). Birds fed the PKM0 and PKM16 diets showed similar feed intake. Similar to weight gain, inclusion of 24% PKM resulted in the lowest (p<0.05) feed intake. Neither the main effect of enzyme addition nor the PKM inclusion×enzyme addition interaction was significant for feed intake.
A significant (p<0.01) interaction between PKM inclusion and enzyme addition was detected for FCR of birds. In PKM0 and PKM8 diets, enzyme addition significantly (p<0.05) improved feed efficiency; whereas, enzyme addition had no effect on feed per gain of birds fed the diets with PKM inclusion above 8.0%.
There was a significant (p<0.001) effect of PKM inclusion on excreta score and excreta DM (Table 2). As the PKM level was increased, the excreta DM increased and excreta score decreased. Neither the main effect of enzyme addition nor the interaction between PKM inclusion and enzyme addition was significant for excreta score and DM.
Nutrient digestibility
A significant (p<0.05) PKM inclusion×enzyme addition interaction was observed for the CAID of N and GE (Table 3). In PKM0 and PKM16 diets, enzyme addition significantly (p<0.05) increased N and GE digestibility but had no effect in the PKM8 and PKM24 diets.

The main effect of PKM inclusion was significant (p<0.001) for the CAID of starch and fat. Inclusion of PKM into the basal diet, irrespective of inclusion level, enhanced (p<0.05) starch and fat digestibility. Whereas the PKM-containing diets resulted in similar starch digestibility, the PKM24 diet had the highest CAID of fat. Neither the main effect of enzyme addition nor the interaction between PKM inclusion and enzyme addition was significant for the CAID of starch and fat.
Inclusion of PKM at 16% and 24% resulted in similar CAID of NDF but higher (p<0.05) than the PKM0 and PKM8 diets. Enzyme addition, regardless of level of PKM inclusion, significantly (p<0.05) increased CAID of NDF.
Nutrient retention and energy utilization
Significant PKM inclusion×enzyme addition interaction was observed for TTR of DM (p<0.05) and NDF (p<0.001; Table 4). Enzyme inclusion had no effect on TTR of DM and NDF in the PKM8 and PKM24 diets, but improved DM and NDF retention in the PKM0 and PKM16 diets.

The main effect of PKM inclusion was significant for the TTR of N (p<0.001) and fat (p<0.01). Diets with no PKM and 8.0% PKM resulted in higher N retention compared to those with 16% and 24% PKM. Inclusion of 24% PKM resulted in the lowest N retention. Birds fed the PKM8 diets had higher (p<0.05) fat retention compared to other PKM inclusion levels.
Increasing the inclusion level of PKM significantly (p<0.001) decreased the TTR of GE. Enzyme addition enhanced (p<0.05) GE retention.
Pam kernel meal inclusion influenced AMEn (p<0.05). There was a significant decrease in AMEn with PKM inclusion of 24%. There was no effect of enzyme addition on energy utilization.
DISCUSSION
Unexpectedly, feeding the PKM8 and PKM16 diets increased weight gain of broilers. Ezieshi and Olomu (2008) reported impaired body weight and feed efficiency in a broiler study with 30% and 32.5% PKM inclusion in starter and finisher diets, respectively. Mardhati et al. (2011) also reported that weight gain and feed efficiency of broilers were poorer in diet with 20% PKM compared to a corn-based control diet. Reduced body weight and feed efficiency have also been reported with PKM inclusion levels above 10% (Soltan, 2009). In most of the studies investigating the effect of PKM inclusion in broiler diets, PKM has been included into a control diet either at the expense of an ingredient (usually the major cereal) in the diet with the consequence of decreased nutrient density or without considering the AA digestibility of PKM. In all the above-mentioned studies, the digestibility of the protein and AA in PKM was not considered and the diets were formulated based on the total protein and AA contents and not the digestible contents. Palm kernel meal has been reported to have a poor protein digestibility (Bryden et al., 2009, CAID of 0.54; Abdollahi et al., 2015, standardized ileal digestibility coefficient of 0.42 to 0.46); which implies that formulating PKM-containing diets based on total crude protein content will fail to meet the birds’ requirement for a balanced diet and impair the growth performance of broilers. Use of digestible AA contents of PKM when formulating broiler diets will eliminate this error and hold a promise to increase the inclusion levels of PKM in practical poultry diets. The PKM sample used in the current study had been previously assessed for the AME and digestible AA contents (Abdollahi et al., 2015), and the data was used to formulate the experimental diets based on apparent digestible AA content. It must be noted that both digestible AA systems, apparent or standardized, are superior to the total AA system. The choice of the appropriate system of digestible AA depends on the method of formulating diets, with CAID being suggested as the most appropriate system to use when the diets are formulated to least-cost (Bryden et al., 2009).
Birds fed the PKM8 diets consumed more feed than other dietary treatments. In agreement to the current study, Adrizal et al. (2011) reported an increased feed intake with increasing PKM level from 0% to 15% and 30% in laying hens. The higher feed intake in birds fed PKM-containing diets has also been reported by other researchers (Onifade and Babatunde, 1998; Sundu et al., 2005; Ezieshi and Olomu, 2008). This observation was attributed to its faster passage rate in the digestive tract (Onifade and Babatunde, 1998), high bulk density and low water holding capacity (Sundu et al., 2006). Onifade and Babatunde (1998) speculated that the higher feed intake in birds fed high levels of PKM in diets was an attempt by the bird to compensate the dilution of the diets and to consume enough nutrients to achieve comparable growth.
Inclusion of 24% PKM in the current study, however, was associated with a marked depression in weight gain, a finding which does not support the implication from several researchers (Panigrahi and Powell, 1991; Perez et al., 2000; Sundu et al., 2006; Adrizal et al., 2011) reporting that PKM can be included in poultry diets up to 40% without impairing performance parameters. Weight gain deterioration of birds fed PKM24 diets although corresponded with the lowest TTR of DM, N and GE, and AMEn, might be explained largely by the feed intake depression at this PKM level. Pellet durability was found to deteriorate with increasing dietary PKM inclusions. This is mostly evidenced by the lowest PDI value determined in the PKM24 diet. Deteriorated pellet quality is also likely to be responsible, in part, for the lowest feed intake and weight gain observed with feeding PKM24 diets.
The present data demonstrated a weight gain response to dietary enzyme addition, even in the PKM0 diet. Enzyme supplementation improved feed efficiency at inclusion levels of 0% and 8% PKM. Although viscosity is not a major nutritional obstacle in corn-based diets, the advantages of exogenous NSP-degrading enzymes on nutrient and energy utilization and broiler performance in corn-based diets have been previously reported (Zanella et al., 1999; Kiarie et al., 2014). In a recent study, Kiarie et al. (2014) reported that xylanase supplementation improved growth performance, nutrient and energy utilization in both wheat- and corn-based diets. The release of previously entrapped nutrients, and not the viscosity reduction, due to hydrolysis of structural arabinoxylans accounted for the positive response to xylanase addition in corn-based diets. These researchers suggested that young birds are sensitive to both soluble and insoluble NSP and could benefit from NSP-degrading enzymes independent of the source of cereal in the diets. In agreement with the current study, Williams et al. (2014) found that supplementation of a corn-soy diet with β-mannanase and a carbohydrase cocktail, separately and intermittently, improved body weight and feed efficiency in broilers. These researchers speculated that the application of a cocktail carbohydrase in corn-soy diets might be the most effective strategy to eliminate the negative effects of various NSP components and to improve the nutrient availability in these conventional feedstuffs.
The beneficial effects of supplementing the poultry diets with β-mannanase have been shown in several studies (Lee et al., 2003; Daskiran et al., 2004; Jackson et al., 2004; Zou et al., 2006; Williams et al., 2014). Lee et al. (2003) reported that supplementation of β-mannanase to corn-based broilers diets containing guar meal reduced intestinal viscosity and enhanced body weight and feed efficiency. Improved feed efficiency as a result of degrading β-mannans through β-mannanase supplementation in a corn-soy-based broiler diet has also been reported by Daskiran et al. (2004). Jackson et al. (2004) reported an improved weight gain and feed efficiency in broilers fed corn-soybean meal based diets supplemented with β-mannanase. The positive growth responses might be, partly, explained by a reduction in energy-draining immune stimulation associated with a decrease in the concentration of β-mannan in the digestive tract of bird. In the current study, calculated β-mannan contents of the PKM0, PKM8, PKM16, and PKM24 diets were 4.50, 29.9, 55.2, and 80.5 g/kg, respectively. It has been shown that β-mannans are components of the surface of many pathogens and perceived by animals as a pathogen-associated antigens stimulating the innate immune system of animals (Jackson et al., 2004; Hsiao et al., 2006; Williams et al., 2014). This immune response, termed as feed-induced immune response, deteriorates animal performance by diverting the energy from growth toward an energy-consuming immune response (Hsiao et al., 2006; Williams et al., 2014). It has been suggested that the inclusion of exogenous β-mannanase hydrolyses the high molecular weight β-mannans into mannose-oligosaccharides which are not recognisable by immune systems. By reducing the immune responses induced by feed, β-mannanase has been shown to spare the energy for production (Williams et al., 2014).
The fact that enzyme supplementation had no effect on the FCR of birds fed PKM16 and PKM24 diets is probably due, in large part, to the fact that the inclusion of PKM at these levels significantly improved feed efficiency compared to the PKM0 and PKM8 diets with no enzyme supplementation. Moreover, there is possibility that β-mannanase inclusion in diets with PKM levels above 8% was not sufficient to hydrolyse β-1,4-glucosidic linkages in β-mannan and to generate performance responses. Daskiran et al. (2004), in a broiler study with diets varying in β-mannan contents, found that diets with higher β-mannan content require higher levels of endo-β-D-mannanase supplementation. These researchers suggested that when ingredients high in β-mannan, such as PKM, copra meal or guar gum, are included in broiler diets, both the dietary enzyme and β-mannan levels should be monitored.
Enzyme supplementation increased the CAID of NDF and TTR of GE. Surprisingly, enzyme addition enhanced CAID of N and GE, and TTR of DM and NDF only in PKM0 and PKM16 diets and not in PKM8 and PKM24; a finding which is not readily explainable. Degradation of NSP by the enzyme cocktail, containing β-mannanase, xylanase, amylase, protease, cellulase and glucanase, might partially explain the positive effect of enzyme addition on nutrient utilization. Corn and soybean meal, though the most common feedstuffs in poultry diets, contain considerable amount of NSP. While corn contains a negligible amount of soluble NSP, it has been reported to have a concentration of insoluble NSP of 80 g/kg which consists mainly of arabinoxylans and β-glucans (Choct, 2006). Ravindran et al. (2014) reported average insoluble and soluble NSP contents of 171 and 16 g/kg, respectively, in soybean meal. A typical corn-soy diet has been reported to contain 123 g NSP/kg, with 111 and 12 g/kg of the NSP being in the form of insoluble and soluble NSP, respectively (Abdollahi et al., 2010). Palm kernel meal sample used in the current study contained 686 and 610 g/kg of NDF and total NSP, respectively (Abdollahi et al., 2015). Saenphoom et al. (2011; 2013) reported that pre-treating palm kernel expeller with exogenous enzyme reduced hemicellulose and cellulose contents by 26% and 33%, respectively. Olaniyi (2014) reported significant reductions in fiber content of PKM as a result of hydrolysing PKM with β-mannanase at 30°C for 60 h. It is also possible that NSP-degrading enzymes by releasing the β-mannans encapsulated in intact cell walls, which can be then depolymerised by β-mannanase, might synergize with β-mannanase as shown by Williams et al. (2014). Aya et al. (2013), using a combination of enzymes containing β-mannanase, α-galactosidase, xylanase, cellulase and glucanase activity, reported higher nutrient (including fiber) retention in enzyme-supplemented diets independent of PKM inclusion level (10%, 20%, 30%, and 40%); an improvement which contributed to the higher weight gain and feed efficiency.
CONCLUSIONS
The present data are suggestive of the potential for PKM to be included in broiler diets. If PKM is to be used in broiler diets, formulating diets based on digestible AA contents and enzyme supplementation are strategies to support optimum growth performance of broilers. When PKM-containing diets are balanced on a digestible AA basis, PKM can be included in broiler diets up to 16% with no deleterious effects on growth performance. Further studies are warranted to elucidate the effects of higher β-mannanase activity, individually or in combination, with NSP-degrading enzymes in broiler diets containing PKM.
ACKNOWLEDGMENTS
We would like to gratefully acknowledge the financial support of the AsiaPac (Dongguan) Biotechnology Co. Ltd. (Guangdong, China) under project APD13001 “Use of β-mannanase in enzyme complexes to enhance the utilization of PKM in poultry diets”.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.