Genetic Relationship of Productive Life, Production and Type Traits of Korean Holsteins at Early Lactations
Article information
Abstract
The present study was performed to study the genetic relationship of productive life with production and type traits of Korean Holsteins at first three lactations. The data for the analysis from 56,054, 28,997, and 11,816 animals of first, second and third parity cows which were born from 2006 to 2011 were collected by Dairy Cattle Improvement Center, National Agricultural Co-operative Federation. Milk, protein and fat yields adjusted for 305 days and average somatic cell score considered as production traits and analyzed type traits were stature, strength, body depth, dairy form, rump angle, rump width, rear leg side view, foot angle, front attachment placement, rear attachment height, rear attachment width, udder cleft, udder depth, front teat placement and front teat length. A multi trait genetic analysis was performed using Wombat program with restricted maximum likelihood animal model composed of fixed effect of birth year, farm and the random effect of animal and random residual effect according to the traits. Heritability estimates of productive life were between 0.06 and 0.13. Genetic and phenotypic correlations between production and productive life traits ranged from 0.35 to 0.04 for milk, 0.16 to 0.05 for protein and 0.18 to 0.02 f 15-0034 (2nd) 150520 or fat. Somatic cells score showed a negative genetic and phenotypic correlation with productive life and also udder type traits, indicating that the selection for higher udder traits will likely to improve resistance to mastitis and persistence in the herd. Among all dairy form type traits, udder characters such as udder cleft showed a significant relationship with productive life. However, a specific change of heritabilities or correlations were not observed with the change of parity. Moreover, further studies are needed to further confirm the significance of the above traits and the effect of parity on above relationships in order to minimize both voluntary and involuntary culling rates while improving herd health and maintaining high yielding dairy cows.
INTRODUCTION
Genetic parameter estimation of economically important traits in dairy cows is essential for early prediction of selection response for achieving goals in efficient breeding programs. Dairy cow production and productive life are most important traits responsible for the profitability of dairy industry.
Milk yield is a very important part of the profitability equation (Luis et al., 2013). High producing herds were profitable in good and poor economies irrespective of herd size. Presently, milk pricing in the world focuses on a multiple component pricing system. As a result, in addition to being indicators of cow health and nutrition, milk component levels have taken on new importance in herd management and now directly impact farm income.
Productive life refers to the total days from the first calving date to the last (culling) date including dry periods (Tsuruta et al., 2005). Extended herd life is economically beneficial due to lower heifer replacement costs and a higher proportion of cows producing at a mature age (Van Arendonk, 1991). Consideration of productive life in selection programs, in addition to economic traits such as milk yield, has increased because of a deterioration of herd life due to intensive selection for enhanced productivity. However, it is difficult to achieve genetic improvement of herd life due to its low heritability (Daliri et al., 2008) and long generation interval. Therefore, researches focus on an alternative method for direct evaluation of sires for high milk production and herd life by using indirect predictors from genetically correlated traits that can be easily measured earlier in the life and are more heritable.
Linear type traits describe biological extremes for a range of visual characteristics of an animal. Linear type traits are relatively easy to measure, and such information is generally available from a cow’s first lactation. In this context, type traits have a direct influence on the management of cows and are related to the profitability of the herd suggesting the importance of studies aimed at reducing involuntary culling and selection for resistant animals. Thus, the objective of the present study was to study the genetic relationship of productive life with production and type traits of Korean Holsteins at first three lactations.
MATERIALS AND METHODS
Data
The data used in the analysis were collected by the milk recording system of the Dairy Cattle Improvement Center, National Agricultural Co-operative Federation for Korean Holstein dairy cattle. Production traits recorded from 723,740 animals and type traits recorded from 349,886 animals were used and, data of production traits, productive life and type traits of first second and third parity animals were collected separately for the analysis. After making three data sets for each parity, preliminary restrictions and eliminations were done by selecting animals born from 2006 to 2011, age at 1st calving between 600 and 800 days. Finally all the recorded traits were pruned to retain the IQR of the data set. Then the data of 56,055 first parity, 28,997 second parity and 11,816 third parity animals were finally used for the statistical analysis. The distribution (basic statistics) of records are shown in Table 1.

Distribution of records used in the analysis according to parity and birth year
Analyzed traits
Dairy cattle Production traits, Type traits and Productive life trait recorded in first, second and third parity separately were considered in the present analysis. Production traits such as 305-day milk yield (Milk305), 305-day protein yield (Prot305) and 305-day fat yield (Fat305) of separate parities were collected. Moreover, somatic cells score (SCS) at first, second and third lactation were also gathered. Productive life was determined by taking the total days from the first calving date to the last (culling) date including dry periods (Tsuruta et al., 2005). Moreover the productive life here was considered similarly for all the three lactations irrespective of parity.
Considered type traits were stature, strength, body depth, dairy form, rump angle, rump width, rear leg side view, foot angle, fore udder attachment, rear udder height, rear udder width, udder cleft, udder depth, front teat placement, and front teat length.
Statistical analysis
Data handling and simple statistics were done by a spread sheet (MS Excel) and a statistical program (SAS, Cary, NC, USA). Genetic parameters and breeding values were analyzed using the restricted maximum likelihood procedure based on an animal model, using the Wombat program (Meyer, 2010). The statistical model for the traits was as follows.
yijkl = observation of considered trait
Herd = fixed effect of herd
BirthYear = fixed effect of birth year
animal = random effect of animal and
residual = random residual effect
If ‘a’ represents the vectors of animal genetic effects for traits, and ‘e’ represents the vector of residual effects, then the random effects can be assumed to follow a normal distribution with zero mean and the following distribution parameters:
Where ‘A’ is the numerator relationship matrix; and ‘G’ is a matrix covariance matrix of the genetic effect between traits. Matrix I is the identity matrix of appropriate dimension in each case, and ‘R’ is the covariance matrix of residual effect.
RESULTS AND DISCUSSION
Heritability (h2) measures the strength of the relationship between phenotype and genotype of an individual animal and it applies to a specific trait measured in a specific population of animals at a specific point in time, if the same trait is measured in a slightly different way in some other group of animals, the estimate of heritability can be expected to be different (Bennet Cassel, 2009).
Heritability estimates for production traits, productive life and SCS are shown in the Table 2. Heritability estimates for productive life were 0.06, 0.13, and 0.11 for first, second and third parities, respectively. A number of publications reported the heritability of productive life varying from 0.04 to 0.17 (Vollema and Groen, 1996; Buenger et al., 2001; Tsuruta et al., 2005; Zavadilová et al., 2009). Heritability estimates of SCS reflect low values and decreased with the increase of parities in the present study as 0.22, 0.19, and 0.11 in first parity to third. Several authors have reported estimates mostly in the range of 0.09 to 0.25 (Banos and Shook, 1990; Castillo-Juarez et al., 2000; Weller and Ezra, 2004; de Haas et al., 2008). Dube et al. (2008) reported heritability for SCS in first, second and third lactation as 0.19, 0.17, 0.19 respectively for South African Holsteins.
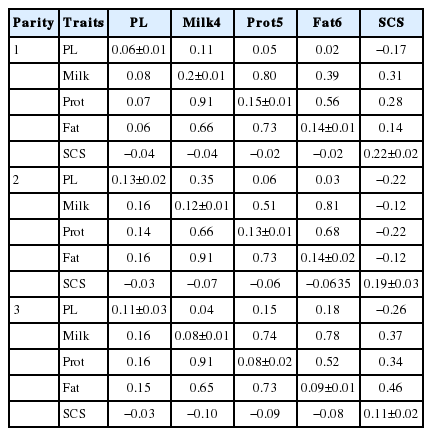
Genetic and phenotypic correlation of productive life and production traits in three parities varied from 0.04 to 0.35 and 0.08 to 0.16 (milk), 0.05 to 0.15 and 0.07 to 0.16 (protein), and 0.02 to 0.18 and 0.05 to 0.16 (fat), respectively. Tsuruta et al. (2004) reported minimum and maximum genetic correlations among milk, fat and protein yields and productive life, in 15 years estimated with a multiple trait random regression model as −0.03 to 0.26 (milk), −0.17 to 0.32 (fat), 0.00 to 0.22 (protein). Further, they pointed out that genetic correlations among traits can increase or decrease over time and even can even change between positive and negative signs. Phenotypic correlations between somatic cell count and yield traits were negative for first, second and third lactations and became increasingly negative as parity increased. However, the genetic correlation was positive in first and third lactations and negative in second lactation. Changes in heritability and correlations are obvious since the parity can influence the pattern of changes in metabolic hormones (Wathes et al., 2007). Moreover, the positive genetic correlations suggest some genetic antagonism between desired increased milk yield and reduced somatic cell count. Cranford and Pearson (2001) found significant, unfavorable correlations between sire predicted transmitting ability for SCS and the number of lactations, total (lifetime) days in milk, and length of productive life of their daughters. We found that a negative correlation of SCS with productive life trait and the value of negative correlation increased with the parity. It suggests that the productive life of dairy cows can be influenced by SCS by the culling of clinically affected animals, as well as the subclinical animals with high SCS.
Animals in the data base that we have used have type scored in their different parities but, only once in their lifetime. On such a situation, one animal can have been scored in the first parity while another animal was scored in the third parity when it was more physically sound than it was in the first parity. Therefore we wanted to observe whether there is an effect on the calculated relationships of productive life with type traits collected in different lactations.
Genetic and phenotypic correlation of type traits and productive life at first, second and third lactations are given in the Table 3. According to the results, body size related traits such as stature, strength, and body depth showed a negative genetic correlation in first and second lactations. Genetic correlation of third lactation and phenotypic correlations of three lactations exhibited a very low positive estimate, close to zero. Dairy form showed moderate negative genetic correlation with productive life as −0.28, −0.22, −0.23 in three parities respectively. Moreover, Lawlor et al. (2005) also found correlation results for dairy form and three different measures of productive life as −0.25, −0.12, and −0.08 for the data received from the Holstein Association of USA. Moreover, they mentioned that the selection against dairy form is the better choice, as it is more strongly associated with fertility and longevity. Rump traits for angle and width also showed a very low negative correlation in both phenotypic and genotypic correlation in first and second lactations. In the third lactation, while the sign changed to positive the value remained close to zero. Similar results were found for body size related traits and rump characters by Cruickshank et al. (2002) for Guernsey Dairy Cattle, Short et al. (1998) for Holstein dairy cattle, Zavadilová et al. (2009) for Czech Fleckvieh cows and Tsuruta et al. (2005) for US Holsteins. In fact, these results reflect that for longer lasting cows for now and in the future the focus should be on more moderately sized animals. Rear leg side view and foot angle also slightly correlate with productive life showing genetic and phenotypic correlations range varying from 0.06 to 0.17 and −0.02 to −0.03, respectively.
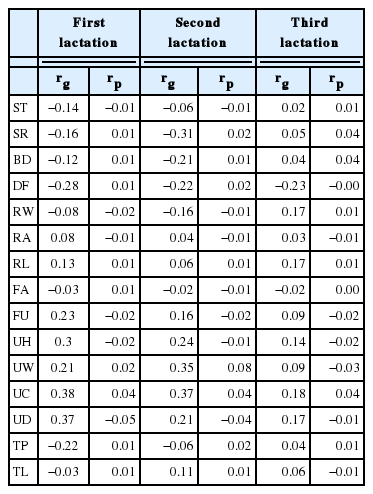
Genetic and phenotypic correlations of productive life and type traits at first three lactations
The conformation of udder and mammary system has a significant impact in the teat lesions, exposure to pathogenic bacteria and milk quality, therefore it directly linked to the udder health. In the present study, the udder and mammary system related traits such as fore udder attachment, udder height, udder width, udder cleft and udder depth showed moderately positive genetic correlation (0.16 to 0.38) in first and second lactations with productive life. In the third lactation it became low (0.09 to 0.18) compared to first two parities. This may be due to smaller number of animal data in the analysis. A moderate genetic correlation for udder and productive life was reported by both Tsurta et al. (2005) and Short and Lowlor (1992) for US Hosteins. Nevertheless, Zavadilová (2009) and Cruickshank et al. (2002) reported a genetic correlation range varying from −0.02 to 0.28 and −0.13 to 0.2, respectively. Udder cleft showed the highest correlation (0.38, 0.37, and 0.18) with productive life in three parities suggesting that stronger cleft translated into a longer stay in the herd by minimizing the potential for injuries and maximizing milking management by keeping the teats in place and udder elevated. In contrast, all phenotypic correlations of udder traits were very low and negative close to zero with the productive life. Furthermore, Short and Lawlor (1992) experienced a low phenotypic correlation compared to genetic correlation with udder traits and productive life. However, the positive genetic correlations of udder traits and productive life in the present study indicate that the animals with higher udder score have an advantage in productive life by high milk production and reduced risk of injury. Teat placement and size are helpful to the ease of milking and susceptibility to injury. In the present study, Teat characters such as teat placement and teat length showed a low negative to positive genetic correlation with productive life range varying from −0.22 to 0.11. Phenotypic correlation of above teat character traits and productive life traits also showed a very low correlation close to zero and the positive correlation became high in the second lactation while negative values were high in first parity correlations. Cruickshank et al. (2002) found a genetic and phenotypic correlation range between −0.25 to −0.03. Moreover, Bouška et al. (2006) reported a negative Pearson correlation coefficient for teat placement (−0.06) and a positive one for teat length (0.06) with uncorrected longevity.
Genetic and phenotypic correlations of milk production traits and type traits are shown in Table 4 and 5. The range of genetic and phenotypic correlations between type trait and milk yield was from 0.74 (dairy form) to −0.23 (rear legs side view) and from 0.75 (dairy form) to −0.19 (udder depth), type traits and protein yield was from 0.75 (dairy form) to −0.46 (foot angle) and from 0.75 (dairy form) to −0.19 (udder depth) and type traits and fat yield was from 0.67 (dairy form) to −0.28 (rear legs side view) and 0.38 (udder width) to −0.25 (udder depth), respectively. Tapki and Guzey (2013) found genetic correlations ranges for type traits and milk yield, from −0.46 (udder depth) to 0.42 (dairy form), protein yield from −0.45 (udder depth) to 0.45 (dairy form) and fat yield from −0.41 (udder depth) to 0.42 (rear teat position), respectively. Lawlor et al. (2005) found an even much higher genetic correlation for milk, protein and fat yields with dairy form. The correlations increase when the cow reaches the third parity in the present study. SCS showed genetic and phenotypic correlations as 0.35 (dairy form) to −0.37 (udder depth) and 0.35 (dairy form) −0.25 (strength) with type traits. Moreover, udder traits are negatively correlated with SCS.
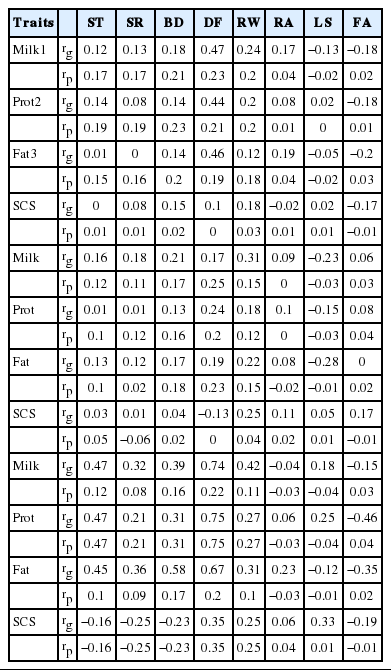
Genetic and phenotypic correlations of milk production traits and type traits
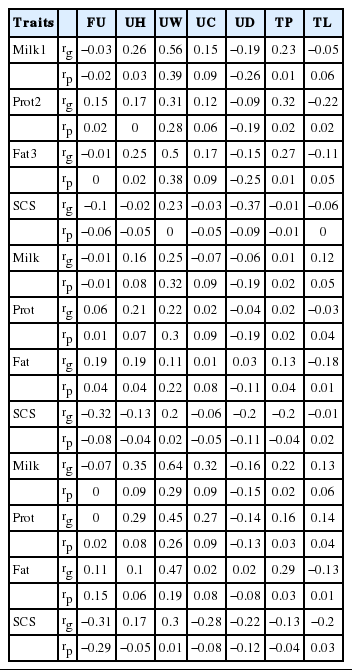
Genetic and phenotypic correlations of milk production traits and type traits
CONCLUSION
Our study resulted in negative genetic correlations with body size traits suggesting that to obtain longer lasting cows for now and in the future selection should focus on more moderately sized animals. It confirmed that the selection against dairy form is the better choice as it more strongly associated with productive life. Comparatively, high genetic correlation of udder traits were observed especially for udder cleft suggest that a strong cleft indicates a longer stay in the herd by minimizing the potential for injuries and maximizing milking management by keeping the teats in place and udder elevated. The animals with higher udder score have the advantage in productive life due to high milk production and reduced risk of injury. The SCS showed a negative genetic correlation with udder trait and productive life, indicating that the selection for higher udder traits will likely to improve resistance to mastitis and persistence in the herd. Specific changes of heritabilities or correlations were not observed with parity. Moreover, further studies are needed to determine the significance of above traits and the effect of parity on above relationships in order to minimize both voluntary and involuntary culling rates while improving herd health and maintaining high yielding dairy cows.
ACKNOWLEDGMENTS
This work was carried out with the support of “Development of systems and techniques for improving resistance and early diagnosis on ketosis in dairy cattle. (Project No PJ010465)” project of the National Institute of Animal Science, RDA, Korea.