




INTRODUCTION
Carcass traits related to beef quality are normally controlled or regulated by a number of genes and single nucleotide polymorphisms (SNPs) in the genes may be significant markers for improved cattle performance (Dekkers and Hospital, 2002; Shin and Chung, 2007). Beef quality and carcass traits usually have low or moderate heritability and are often recorded post-slaughter (Marshall, 1999; Gill et al., 2009), therefore, SNPs can be used as markers for indirectly improving beef quality traits instead of direct measurements (Ge et al., 2003; Allan et al., 2007).
During the last decade two promising SNPs have been identified and are being used in selection programs of the economically important traits in the livestock industry. One of the SNPs is in the calpain 1 gene (CAPN1, (mu/I) large subunit), which is known to associate with meat tenderness (Smith et al., 2000; White et al., 2005; Casas et al., 2006). The other SNP in the 5′-AMP-activated protein kinase subunit gamma-3 gene (PRKAG3), which affects high glycogen content in porcine skeletal muscle (Li et al., 2012; Ryan et al., 2012).
DNA methylation mediates the interaction of cattle with environment and nutrition (Liu et al., 2012), thus it is important to study the genetic effects of SNPs in epigenetic-related genes in beef quality and carcass traits. Recent studies by our research group fellows (Guo et al., 2012; Liu et al., 2012) suggested that epigenetic related genes (DNA (cytosine-5-)-methyltransferase 3 alpha [DNMT3a], DNA (cytosine-5-)-methyltransferase 3 beta [DNMT3b]) play a role in meat quality and carcass traits. The SNPs in DNMT3b were found significantly associated with meat quality and carcass traits (Liu et al., 2012; McClure et al., 2012). The present study continues our previous study (Liu et al., 2012), which was focused on few SNPs in DNMT3b gene with beef quality traits.
The objectives of this study were to analyze the effects of SNPs on fourteen beef quality and carcass traits in seven epigenetic-related genes (four DNA methyltransferases genes, namely DNMT1, DNMT3a, DNMT3b, DNMT3L; two argonaute genes argonaute RISC catalytic component 1 [Ago1] and Ago2; and histone deacetylase 5 gene [HDAC5]) and the two beef quality candidate genes (CAPN1 and PRKAG3), and to develop potential markers for improvement of meat quality traits in Snow Dragon beef cattle. To provide basic molecular information, we also described the SNPs in other thirteen cattle breeds.
MATERIALS AND METHODS
Samples and meat quality traits
Muscle, semen and ear tissue samples were collected from 383 individuals belonging to 14 breeds of cattle. Amongst these 14 breeds, 6 were native Chinese cattle while the other 8 were exotic breeds (Table 1).
The records of 14 meat quality traits were available for 147 individuals from the Snow Dragon beef population (the offspring of Wagyu and Filial 1 hybrid (F1) crossbred cows of Limousin by Fuzhou yellow cattle, supplied by Xuelong Industrial Group, Dalian, China) and were used for association analysis of genotypes with meat quality traits in the present study. The fourteen meat quality traits include dressing percentage (DP), carcass weight (CW), chilled carcass weight (CCW), rib-eye width (RW), rib-eye length (RL), rib-eye area, flank thickness (FT), chuck short rib thickness (CT), chuck short rib score (CS), back-fat thickness (BFT), chuck flap weight (CF), fat color score, lean meat color score (LCS), and marbling score (MS).
Detection and genotyping of the polymorphisms
Genomic DNA was extracted from the muscle and ear tissue samples using the commercial kit of genomic DNA purification Wizard (Promega Corp., Madison, WI, USA) according to the manufacturer’s instructions, while DNA for semen samples were extracted using high salt extraction procedures (Liu, 2007; Qin, 2008). Primer pairs were designed to identify SNPs which covered all coding regions and intron sequences as well as the 5′ regulatory region 2,000 bp upstream of the transcription initiation site using the software Primer 3 (v.0.4.0) and Oligo6.0. A DNA pool was prepared consisting of ten cattle (50 ng/μL per sample) randomly selected from Limousin, Wagyu, Simmental and Angus.
Polymerase chain reaction (PCR) amplifications for the pooled DNA were performed in a final reaction volume of 25 μL consisting of 50 ng genomic DNA, 0.5 μL each primer, 2.5 μL 10× PCR buffer, 2.5 mM of dNTP and 1 U Taq DNA polymerase (TaKaRa, Dalian, China). The PCR protocol was 5 min at 95°C for initial denaturation followed by 40 cycles at 95°C for 30 s, 59°C for 30 s, 72°C for 30 s and a final extension at 72°C for 10 min for all the primer pairs. Matrix-assisted laser desorption/ionization time of flight mass spectrometry (MALDI-TOF MS, Squeno MassARRAY, Bioyong Technologeies Inc. HK, China) assay was used for individual genotyping of 383 individuals belonging to 14 breeds of cattle.
Statistical analysis
The genotypic and allelic frequencies of the SNPs were calculated using PopGene 3.2. Association analysis between the 15 SNPs and beef quality traits in the population of Snow Dragon beef was carried out using the general linear model procedure of SAS 9.1 with the following fixed effect model:
Where y represents the phenotype records of carcass and meat quality traits; μ is overall mean; gi is the fixed effect of the genotype; sj is the fixed effect of sex; mk is the fixed effect of month-old; dl is the fixed effects of slaughtering date; e is the random error. The statistics were presented as probability values and least squares means±standard error, and a p value of <0.05 was considered statistically significant.
The combination effects of SNP15 with other 14 SNPs on beef quality traits in the Snow Dragon beef population were analyzed with the following model:
Where y, μ, sj, mk, dl, and e are the same as shown in Equation 1, and ci is the effect of the combination genotypes between two SNPs.
RESULTS
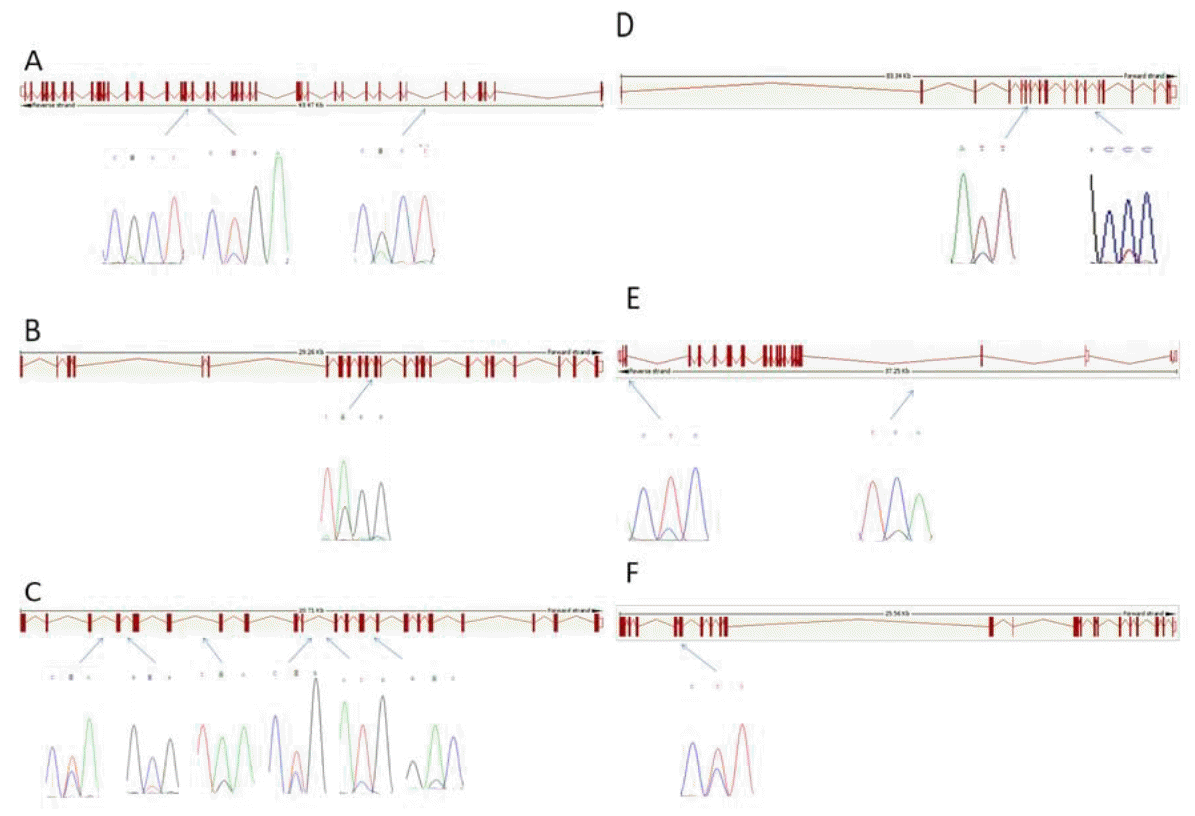
Investigation of the promoter and the exon sequences of pooled DNA revealed 16 SNPs in the 9 genes, amongst these 16 SNPs, 7 were novel (Table 2). A total of 3 SNPs were found in the exon regions including SNP1 (13154420A>G) in exon 17 and SNP2 in exon 18 (13153987T>C) of DNMT1, and SNP15 (45219258C>T) in exon 4 of CAPN1. Whereas, the other 13 SNPs were found in the intron regions (Table 2, Figure 1).
Genotype and allele frequencies
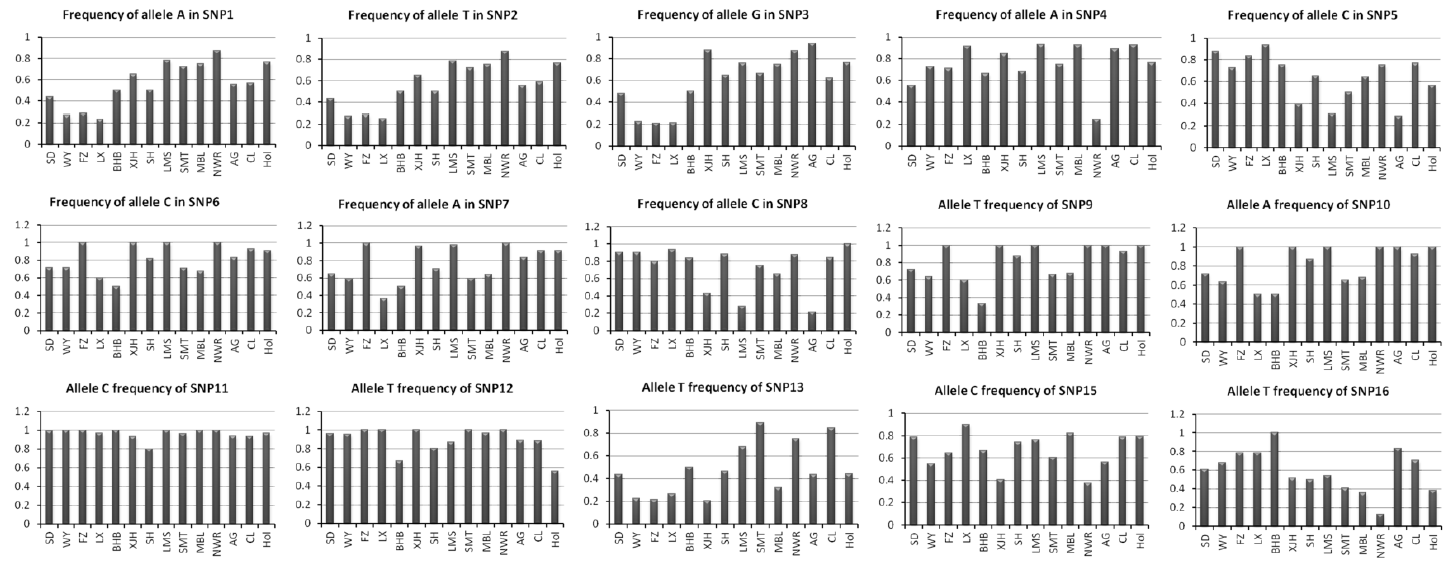
Amongst the 16 SNPs, except SNP14, 15 were successfully genotyped using MassARRAY technique. The Genotype and allele frequencies are shown in Table 3. The wild-type and mutation-type allele frequencies of the 15 SNPs identified in the genes DNMT1, DNMT3a, DNMT3b, Ago2, HDAC5, CAPN1, and PRKAG3 in all 14 breeds of cattle are shown in Figure 2. Generally, the allelic frequencies of wild-type allele were higher than those of the mutation-type allele in each breed, and also there was a plain tendency in the allelic frequency of the native and foreign breeds i.e. the allele T frequencies of SNP2 (13153987T>C) in native breeds (p<0.5 except for Sanhe cattle) were lower (>0.5), whereas, the allele C frequencies of SNP5 (63037313C>T) in native Chinese breeds were higher than those in exotic breeds. The allele and genotype frequencies of the SNPs were very similar between Snow Dragon beef and Wagyu cattle.
Association studies
Out of the 15 SNPs, 8 SNPs in 4 genes (DNMT1, DNMT3a, DNMT3b, and CAPN1) were found significantly associated with carcass and meat quality traits (Table 4). SNP1 (13154420A>G, National Center for Biotechnology Information [NCBI] reference no rs135600607) of DNMT1 was a silent mutation located in exon 17 of chromosome 7 and was found significantly associated with RW and LCS (p<0.05). Animals with mutation genotype GG in SNP1 performed better than the wild type genotype AA for the two traits.
SNP4 (76198537A>G) was found to be a novel SNP and located at intron 11 of DNMT3a gene. Allele A of SNP5 was significantly associated with CF (p<0.05) and highly significantly (p<0.01) associated with CW, CCW, FT, and CT. Compared with the mutation genotype GG, individuals with the wild-type genotype AA showed remarkable increase in CW, CCW, FT, CT, CS, and in CF.
Six SNPs were uncovered in the DNMT3b gene. Of which, five SNPs i.e. SNP5 (63037313C>T), SNP6 (63037426C>T), SNP7 (63030432A>G), SNP9 (63033215T>C) and SNP10 (63039420A>G) were significantly associated with beef quality traits (p<0.05). Individuals with the mutation genotype TT of SNP5 (63037313C>T), compared to wild-type genotype CC, showed an increase of 84.12 kg in CW, 1.93 cm in RL, 0.70 cm in FT, 0.61 kg in CF and a decrease of 1.15 cm in BFT. In SNP6 (63037426C>T) the difference between the wild type CC and mutation type TT genotype was 0.50 kg for CF. The mutation genotype GG of SNP8 showed an increase of 0.58 in CF, whereas, for SNP9 the differences between the wild type genotype TT and the mutation genotype CC for CF and BFT were 1.15 kg and 0.61 cm, respectively. Individuals with the heterozygous genotype (AG) in SNP10 showed an increase in DP, BFT, and MS in contrast with the homozygotes. The rest of the SNPs were significantly (p<0.05) associated with CF except SNP11 (63039420A>G).
The SNP15 (45219258C>T, NCBI reference No rs17872079) of CAPN1 gene is a silent mutation in exon 4 on chromosome 29, which was significantly associated with CT and LCS (p<0.05). Individuals with the wild type CC genotype showed increases of 0.20 cm in CT and 1.00 in LCS.
The effects of combined genotypes on meat quality
The SNP15 (45219258C>T) of CAPN1 was significantly associated with CT and LCS. Therefore, SNP15 (45219258C>T) was selected for analysis in combination with 14 other SNPs to investigate the effects on beef quality traits in the Snow Dragon beef population. Interestingly, all of the 14 combination genotypes were associated with LCS, and the lowest score was showed by combination genotype of NNTT, probably because of the strong association of TT genotype of SNP15 with lower LCS. The results of combination genotypes are mentioned in Table 5.
DISCUSSION
In the global scenario of increasing demand for protein sources and food, improving beef quality traits through marker assisted selection has received increasing attention (Pariacote et al., 1998; Lindholm-Perry et al., 2012; McClure et al., 2012). The use of effective molecular markers in selection for beef quality traits is a worthy approach that can bring improvement in beef quality (Hoey et al., 1995; Zan et al., 2007).
In the present study, fifteen SNPs in seven epigenetic-related genes and two meat quality candidate genes were detected in a Snow Dragon cattle population as well as thirteen other cattle breeds. Results of the study indicated that 8 out of 15 SNPs were significantly associated with carcass and beef quality traits. In these 8 SNPs, SNP1 (13154420A>G) and SNP15 (45219258C>T) were silent mutations found in the exon region of the DNMT1 and CAPN1 genes and were significantly associated with meat quality traits. DNMT3a and DNMT3b were found important genes in which SNPs were significantly associated with a couple of beef quality traits. Moreover, seven SNPs in the 5 genes (DNMT3L, AGO1, AGO2, HDAC5, and PRKAG3) were non-significantly associated with all of the beef carcass and quality traits in the Snow Dragon beef population.
The DNMT1, DNMT3a, and DNMT3b are DNA methylation-related genes and were found significantly associated with carcass traits related to beef quality. Recently, studies had indicated that gene expression of DNMT3a and DNMT3b were related to fat deposition depicting their potential role in adipose tissue (Kamei et al., 2010). Our recent studies on bovine DNMT3a and DNMT3b showed that DNMT3a was more highly expressed in liver and muscle tissues in beef cattle than DNMT3b, and both of the genes were found significantly associated with beef quality traits (Guo et al., 2012; Liu et al., 2012).
In the present study, the SNPs in DNMT1 and DNMT3a gene were significantly associated with many of the beef quality traits. Mutant allele “G” of SNP1 (13154420A>G) and wild type allele “A” of SNP4 (76198537A>G), associated with improvement in beef quality traits, can be used as potential markers for beef quality improvement. DNMT3b gene was revealed to be a noteworthy gene having 5 SNPs significantly associated with many beef quality traits. The significant influence of SNPs in DNMT3b gene on CF revealed that this gene could be a potential candidate gene in future studies for beef quality improvement.
The CAPN1 gene has been extensively studied and it has been reported that SNPs in this gene were significantly associated with carcass and meat quality traits, especially, with meat tenderness and MS in beef cattle (Page et al., 2004; Corva et al., 2007; van Eenennaam et al., 2007; Pinto et al., 2010). In the present study, SNP15 was chosen for combination genotype association with the rest of 14 SNPs to analyze the effects on beef quality traits. Interestingly, the combination genotype effect of SNP15 (45219258C>T) with all 14 other genotypes showed significant association with LCS. Therefore, it is proposed to consider this SNP as a genetic marker for selection of higher meat quality in beef cattle. Moreover, the results indicated that the effects of combination genotypes were higher than the single SNP effect on the corresponding phenotype.
Initially the fixed effects of the genotype, sex, month-old and slaughtering date were used for data analysis in the model. The results revealed that only SNP had significant effect on the studied phenotypes whereas, all other fixed effects were non-significant effect. Considering the reduced model excluding all other fixed effects except the effect of the genotype changed the results but these results are in line with the previous results (the individual SNP effect remained significant on most of the traits under study). We have provided the fresh results as additional files (Supplementary Tables 1 and 2). These results show that the SNPs alone have significant effects on the phenotypes.
In PRKAG3 gene, SNPs are known to be correlated with gene expression and meat quality phenotypes and this gene has been proposed as a positional and functional candidate gene for quantitative trait loci exerting significant effects on the beef quality traits (Rohrer et al., 2012). More than 100 SNPs have been identified in the CAPN1 gene of cattle and out of these, 4 SNPs have revealed significant effects on meat tenderness (Smith et al., 2000; Juszczuket et al., 2004; Pag et al., 2004).
For the fourteen cattle population, we found that most wild type alleles had higher allelic frequencies in the population than the mutation alleles. These findings suggested that wild type alleles were not frequently mutated and maintained differential tendency for the native and imported breeds. The allelic and genotypic frequencies of Wagyu and Snow Dragon beef were in accordance with each other due to their common genetic background (As, Snow Dragon beef are the offspring of Wagyu and F1 crossbred cows of Limousin by Fuzhou yellow cattle). These results were in accordance with our previous study (Liu et al., 2012).
CONCLUSIONS
Amongst 9 screened and analyzed genes in the present study, SNPs in only 4 genes i.e. DNMT1, DNMT3a, DNMT3b, and CAPN1 were found significantly associated with carcass and beef quality traits. In the DNA methylation-related genes, 7 SNPs significantly associated with many of the carcass traits related to beef quality suggesting that these genes could be potential candidate genes for genetic improvement in beef quality traits. Moreover, the significant association of SNP15 in CPAN1 gene alone or in combination with all 14 other SNPs with lean meat score strongly qualify this SNP to be used as a genetic marker in selection for improvement in this particular beef quality trait. The SNPs found in the present study can be used in further studies to investigate the functional effects of these SNPs on meat quality traits.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print







