



INTRODUCTION
Coat color and pigmentation on various body parts of animals are a readily observable visual phenotype and are mostly considered for selection criteria because even small changes in coat color influences the organism’s ability to survive or reproduce (Endler, 1993; Safran and McGraw, 2004) in domestic and wild animals. Becerril et al. (1993) demonstrated that the degree of white spotting in Holstein cattle influenced the level of milk yield and reproductive performance in terms of solar radiation in the environment (Becerril et al., 1993).
All mammalian pigmentation result from the pigment producing cells (melanocytes) and therefore, variations in vertebrate skin and hair color are due to varied amounts of eumelanin (brown/black) and pheomelanin (red/yellow) produced by the melanocytes. Melanocytes originate in the neural crest and migrate to dermis during mammalian development (Jackson, 1994; Klungland and Vage, 2000). Pigmentation biology has been extensively studied in mice and humans to understand its pleiotropic effects (Jackson, 1997). Mammalian pigmentation model systems have served to study molecular, cellular and developmental processes in detail (Silvers, 1979). Till date, over 150 genes known for animal coat color and patterning have been identified and reported (Roulin, 2004; Mundy, 2005; Hoekstra, 2006; Protas, 2008).
Few earlier studies in vertebrate species have revealed the role of melanocortin 1 receptor (MC1R), formerly named by alpha melanocyte stimulating hormone receptor (α-MSHR) in the regulation of eumelanin and pheomelanin pigment synthesis in melanocytes (Robbins et al., 1993).
The Korean Hanwoo cattle exhibit three different coat color phenotypes viz., black, brindle and yellowish-brown and concomitant muzzle color of black, yellowish-brown and black spotted brown, respectively (Seo et al., 2007). Our group previously reported the molecular variation in pigmentation genes (TYRP1 (exon 5) and MC1R genes) in Hanwoo cattle and Black Angus. Sequence variants particularly in MC1R were reported to be associated with total melanin and eumelanin in breeds (Mohanty et al., 2008). Pigmentation genes such as TYR (tyrosinase), TYRP-1 (tyrosinase-related protein 1), DCT (previously TYRP-2 or tyrosinase-related protein 2) and MC1R play a vital role in cattle melanogenesis (Seo et al., 2007). In addition, both α-MSH and nitric oxide were reported to accelerate the production of eumelanogenic melanin (Hunt et al., 1995; Thody and Graham, 1998; Ito et al., 2000) while L-cysteine tunes up the pheomelanin production (Hunt et al., 1995). The present study was designed on the basis of our previous findings (Mohanty et al., 2008) and to further investigate the effect of α-melanocyte-stimulating hormone (α-MSH), nitric oxide and L-cysteine on melanin production and expression of related genes MC1R, Tyr, Tyrp-1 and Tyrp-2 in muzzle melanocytes of Hanwoo breeds (i.e., black, brindle and brown). Earlier workers have studied the diverse patterns of mammalian coat color by the quantity and distribution of two types of organic pigments such as eumelanin (black to brown) and pheomelanin (yellow to red) (Ito, 2003; Barsh, 2006). Both are produced by melanocytes in the hair bulbs and basal epidermis. We herein report that pigmentation in melanocytes is dependent on the melanogenic factors such as α-MSH and nitric oxide which adds eumelanin and pheomelanin to the melanocytes respectively. To the best of our knowledge our group is the first to have studied the effect of abovementioned factors on the production of melanin in muzzle melanocytes of native Korean Hanwoo breeds.
MATERIAL AND METHODS
Sampling and cell culture
Muzzle samples were taken from three native Hanwoo cattle which had the phenotype of black, brindle and brown coat color, respectively. The muzzle samples were collected from the experimental animals of the Chonbuk Livestock Research Station after slaughter with the kind permission of ethical committee of Chonbuk National University. Approximately 2×2 cm muzzle samples were collected from the slaughtered animals. After sampling the tissues were ice-packed supplemented with antibiotics. The collected samples were transferred to the laboratory within 40 min for further processing. The samples were processed for melanocytes isolation by the procedure as described by Ian Freshney (Ian Freshney, 1994) with suitable modifications. Briefly, muzzle samples were washed with sterile D-PBS (Gibco) to remove potential surface contamination. Most of the fat was trimmed off and discarded.
The muzzle tissues were cut into pieces (approximately 5×5 mm) and cultured with 0.25% trypsin solution (Gibco) at 4°C for 24 h and rocked gently every after 1 h. After incubation the fragments were taken into a dry sterile petri-dish with epidermal side down and the dermis was cleaned. Further, the tissues were put into 0.02% EDTA and vortexed gently to disintegrate the cells and centrifuged at 350×g for 5 min. The pellets were re-suspended in melanocyte medium DMEM (Gibco) supplemented with 2.5% fetal bovine serum (FBS, Gibco), 2 mM glutamine, 1 mM sodium pyruvate, 1% non-essential amino acids, 2.5 ng/ml epidermal growth factor, 25 μg/ml bovine pituitary extract, 10 ng/ml Tetra-decanoy1 phorbol acetate (TPA), 100 U/ml penicillin, 100 μg/ml streptomycin, plated into uncoated tissue culture flasks (Falcon, Bacton Dickson Labware, Franklin Lakes, NJ, USA) and incubated in a CO2-regulated incubator (Thermo Electron Corporation, Revco, USA) in humidified 95% air/5% CO2 atmosphere. Bovine pituitary extract (BPE) contains a variety of growth factors and hormones. So BPE was added to supplement the growth factors to the medium. In case of contamination with fibroblasts cells the culture medium was supplemented with geneticin (Sigma, USA) at a concentration of 100 μg/ml for 3 to 4 days. The pure cells were then subcultured. Finally the culture medium was supplemented with 15% horse serum and 2.5% fetal bovine serum to enhance the cell growth. Medium was changed after three days in a week.
Treatment
The second passaged cells (2.5×105) seeded into confocal dishes (Falcon, Bacton Dickson Labware, Franklin Lakes, NJ, USA) were allowed to attach overnight and subjected to three different treatments i.e. α-MSH, nitric oxide donor (sodium nitroprusside SNP) and L-cysteine. The dosage for α-MSH and SNP were 100 nM and 50 μM for each dish respectively. L-cysteine was added as 0.07 mg/ml of media. For α-MSH treatment the media used was without bovine pituitary extract (BPE) and treatment was done after 12 h of culture. For nitric oxide treatment, sodium nitroprusside (SNP) was used as a donor and the cells were treated after 24 h in culture. For L-cysteine, treatment was done after 12 h of culture in Dulbecco’s (cysteine free) media. In controls only medium was replaced with fresh medium after 24 h. All the experiments were done in triplicates along with suitable controls.
Melanin measurements
Total melanin in the cell suspension was measured spectrophotometrically according to procedure described in our previous publication (Mohanty et al., 2008). All the measurements were performed in triplicates. Briefly, 200 μl of cell suspension (4× 105 cells) was taken and to this 2 μl of soluene:water (9:1v/v) mixture was added and mixed properly (~10 s). The mixed samples were placed in a water bath maintained at 80°C for 30 min; the samples were taken out, remixed properly and kept again in the same water bath for further 15 min. Finally, samples were cooled at room temperature. Standards were also processed in the same way. Absorbance was measured at 500 and 650 nm by spectrophotometer using sepia melanin as the standard.
For eumelanin estimation, permanganate oxidation was performed to obtain pyrrole-2, 3, 5-tricarboxylic acid (PTCA) and its quantification by HPLC was in accordance with earlier established procedures (Ito and Fujita, 1985; Mohanty et al., 2008). Briefly, 200 μl cell suspensions (4×105 cells) were taken and to this 1 ml of 1 M H2SO4 was added, the resulting solution was mixed properly. To 0.1 ml of BSA (20 mg/ml in H2O), 15 ml of 3% KMnO4 (0.15 g in 5 ml H2O) was added and mixed properly, purple color disappears. Again, 15 ml of 3% KMnO4 was added and mixed for 10 min. After that 100 ml of 10% Na2SO3 was added. Finally, 7 ml of peroxide free Di-ethyl ether was added and mixed. The organic phase was separated and collected in a separate tube. This step was repeated. The collected organic phase was dried under a steam of nitrogen. To the organic residue, 300 ml de-ionized H2O was added and samples were filtered through 0.45 μm filter. HPLC (LC-20AD, Communication bus module, CBM-20A, Shimadzu, Japan) was performed at 45°C using a pheomenex RP-C18 column (250 mm×4.6 mm, 7 μm particle size) with a flow rate of 0.8 ml/min using isocratic pump. The mobile phase consisted of 0.01 M potassium phosphate buffer (pH 2.1): methanol 98:2 (v/v). 20 μl of sample was injected and PTCA peak was detected at 269 nm. A standard curve was obtained using Sepia melanin (Sigma) and the retention time of the PTCA peak was established by pre-running a PTCA standard. All estimations were performed in triplicate and the average values were used for analysis in this study as described earlier (Mohanty et al., 2008). The PTCA values obtained were multiplied by a factor of 50 to obtain the eumelanin values (Ito and Fujita, 1985).
mRNA expression
Total RNA was extracted from 106 cells of each breed using with Trizol solution (GIBCO-BRL, Burlington, ON, Canada) according to the manufacturer’s specifications (Chomczynski and Sacchi, 1987). First strand cDNA was synthesized from 1 μg of the total RNA using the M-MLV Reverse transcriptase with the anchored oligo d(T) primer. The mRNA expression of selected genes was measured by reverse transcription polymerase chain reaction (RT-PCR). The PCR program initially started with a 94°C denaturation for 4 min, followed by 20 to 42 cycles of 94°C/45 s, Ta°C/45 s, 72°C/1 min. Linear amplification range for each gene was tested on the adjusted cDNA and here primer sequences, optimal PCR annealing temperatures (Ta) and cycle number are omitted. After electrophoresis of PCR products, the gels were digitalized (VersaDoc 3000, Rio-Rad, USA) and relative density was determined against an arbitrary unit (Quantity One, Bio-Rad, v4.62, USA).
Confocal microscopy
Confocal microscopy was performed according to the standard procedure. Briefly, a portion of cells (1×106 cells) was taken and centrifuged to get the pellet. Supernatant was discarded and the pellet was suspended into phosphate-buffered saline (PBS) and gently shaken at room temperature. PBS was removed and cells were re-suspended into the same buffer, this step was repeated twice. Next, the PBS was removed and pellet was transferred into the ethanol (1 ml) at −20°C. The cells were kept in ethanol for 15 to 20 min at −20°C. After 20 min the ethanol was discarded and the cells were rehydrated with PBS at room temperature for 15 min. Finally the PBS was removed and cells were stained with Propidium Iodide. A cover slip was used to distribute the dye evenly on the slide and poly-l-lysine was used for the fixation. Samples were incubated at 37°C for 30 min in dark. The cover slip was removed and the fixed cells were rinse briefly with PBS to remove the unbound dye. Excessive liquid was removed from the slides by gently blotting around the samples with an absorbent tissue. The effect of treatments on the cells was assessed by propidium iodide staining after visualization under confocal microscope (Carl Zeiss, LSM 510 META, Germany) at regular time intervals viz., 24, 48, 72 and 96 h.
Statistical analysis
Least square means of total melanin and eumelanin as a function of breeds in samples were calculated by using a general linear model (SAS, 1997), and the significance was evaluated by a pairwise difference analysis at p<0.05.
RESULTS
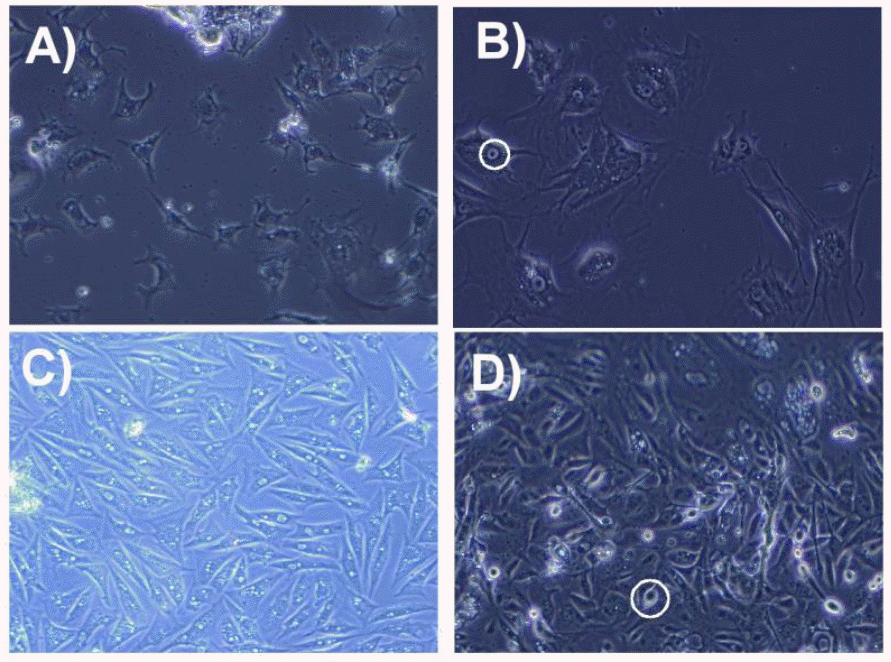
Melanocyte cells (2nd passage) were grown to confluence (~80%) and were then subjected to three different treatments i.e. α-MSH, nitric oxide donor (SNP) and L-cysteine as aforementioned in the in the materials and methods. All the experiments were carried out in triplicates along with suitable controls. The culture protocol that was employed enabled us to establish and maintain melanocytes from Hanwoo muzzle under in vitro conditions up to several months. Morphology of the cultured cells was evaluated throughout the incubation period. Primary cells were maintained in basal medium with 2.5% fetal bovine serum. The primary cells showed dendrite formation, cells connecting each other (Figure 1a). The first passage cells were maintained in 5% serum. At this stage the cells showed maturation, high dendriticity and slight accumulation of melanin in the circles (Figure 1b). Generally the dividing cells showed polygonal to elongated morphology with vacuolation (Figure 1c). At 21 days of incubation, melanocytes showed the accumulation of melanin. Prominent rounded structures in circles (Figure 1d) were observed which might be dead cells. The cell death was likely due to the melanin toxicity or bursting of cells in the culture medium.
Moreover, significant increase in total melanin was also observed with L-cystiene in Hanwoo brown. On the other hand a significant increase was found in eumelanin when melanocytes were supplemented with L-cystiene in Hanwoo brown. Conversely, L-cystiene showed a significant suppression of eumelanin content in Hanwoo black but no significant change was found in Hanwoo brindle. However, a significant increase in eumelanin was found in Hanwoo brown by α-MSH and nitric oxide treatments (Table 1).
The statistical significance of total melanin and eumelanin in muzzle samples was determined (SAS Institute Inc., USA, 1997) and the effect of breed (i.e., coat color) on melanin content is presented in the current study (Table 1). Significant (p<0.05) variation in total melanin and eumelanin content among different phenotypes was observed. A significant increase was found in total melanin content when melanocytes were treated with nitric oxide and α-MSH in Hanwoo black and brindle.
Furthermore, we studied the mRNA expression of MC1R, Tyrp1, Tyrp2 and Try genes in melanocytes treated with L-cysteine, α-MSH and nitric oxide from at least three unrelated Hanwoo phenotypes. Table 2 lists the mRNA expression of MC1R, Tyr, Tyrp1 and Tyrp2 genes in melanocytes culture for black, brindle and brown Hanwoo treated with L-cysteine, α-MSH and nitric oxide, respectively. Semi-quantitative RT-PCR showed the up regulation of Tyr in all selected cattle by nitric oxide and in α-MSH treatments, whereas Tyr, Tyrp-2 and MC1R genes are down regulated in L-cysteine treated melanocytes from all the three phenotypes (Table 2).
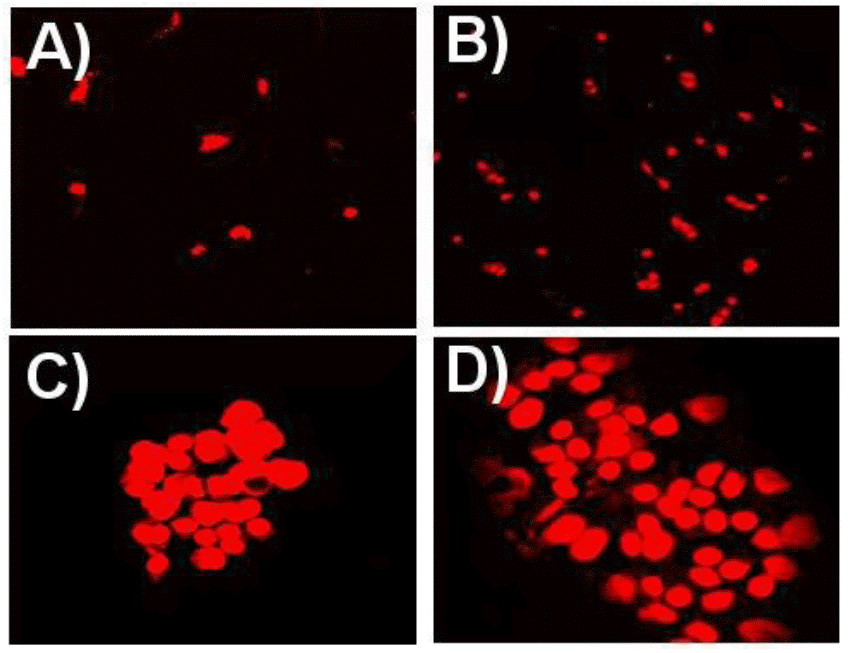
To get more insight into the changes of melanocytes culture, confocal microscopic studies were done. The numbers of dead cell assessed by confocal microscopy after propidium iodide staining (Figure 2) were tabulated (Table 3) up to 96 h of incubation period. Significant increases (p<0.05) were found in the number of dead cells with an increase in incubation time in both the groups. A significant increase was observed in dead cells from 24 to 72 h when cells were treated with nitric oxide and α-MSH respectively, however, with L-cysteine the values were non significant. In the present study, the highest number of dead cells was obtained when the melanocyte culture was supplemented with α-MSH (Figure 2) and also after 96 h in all treated cultures.
DISCUSSION
Earlier reports on melanocytes and melanoma cells in culture mainly described their typical structure and metabolic features (Bucek and Vrba, 1976; Bhavanandan, 1981; Waldrep and Kaplan, 1986; Hu et al., 1987). Korean Hanwoo cattle exhibit three different coat colors and concomitant muzzle colors (Seo et al., 2007; Mohanty et al., 2008) but the behavior of melogenic factors on Hanwoo muzzle melanocytes were never before investigated in culture. We therefore, investigated the effect of α-MSH, nitric oxide and L-cysteine on them.
Most of the protocols used for the various melanocyte cultures (Bucek and Vrba, 1976; Bhavanandan, 1981; Waldrep and Kaplan, 1986; Hu et al., 1987; Mohanty et al., 2008) involved the use of mitogens such as cholera toxin. In this study, melanocytes were grown without addition of cholera toxin. Cholera toxin mimics melanocyte stimulating hormone in inducing differentiation in melanoma cells and alter morphology of melanoma cells in culture. To avoid toxin related problems, melanocytes were grown without the supplementation of cholera toxin but the growth rate was very slow. To obtain primary cultures enriched with melanocytes, most of the dermis was removed from the muzzle specimens; this minimizes fibroblast contamination. An overnight trypsinization was carried out at room temperature to enhance the melanocyte ratio. For better assessment of differentiation, only fully differentiated melanocytes were used in the present study. Selective differentiation of melanocytes was obtained by using trypsin (0.25%). Hanwoo muzzle tissues were cultured with trypsin (0.25%) solution at 4°C for 24 h. Confluent primary cultures were submitted to trypsin treatment and passaged. If residual keratinocytes were still observed after this passage, a further trypsinization was carried out. In the present study trypsin (0.25%) produced the best results. Moreover, the advantage of the trypsin digestion method is that it is a relatively simple method and easy to perform. Also geneticin effectively eliminated contaminant cells, mainly fibroblasts cells, without toxic effects on melanocyte culture. So the method used in this study provided pure culture with good growth and melanogenic capacities in vitro.
In mammals, melanogenesis is highly regulated and complex phenomenon occurring within specialized cells termed melanocytes. The melanin synthesis in the melanocytes is under genetic control. This control is also maintained when the cells are removed from their natural environment and grown in culture. Many studies of melanogenesis by epidermal melanocytes have been reported and it is well established that epidermal melanocytes do produce melanin in vivo and in vitro. Hu et al. (1995) reported that the culture system is capable of generating large numbers of human uveal melanocytes cells in pure culture. For an accurate investigation of melanogenes in vitro, it is essential to generate a large population of melanocytes in pure culture. Study of melanogenesis by uveal melanocytes in vitro has been hampered by their low proliferative potential and the tendency for contamination with other cell types under usual culture conditions. Melanin content and the production of melanin by uveal melanocytes in vitro apparently are influenced by racial and genetic factors because uveal melanocytes from a black donor showed significantly higher melanin content, rate of melanin production, and tyrosinase activity, as well as larger melanosomal size, than seen in uveal melanocytes derived from white donors. The content of melanin per cell in cultured uveal melanocytes is not only determined by the production of melanin but also by growth rate. In stationary cells, the melanin produced accumulates within the cell and results in a rapid increase of melanin content per cell. In growing cells, the melanin is diluted to daughter cells during division. If the melanin production rate equals the rate of dilution, the melanin content per cell would remain unchanged. In rapidly growing cells, if the dilution rate is greater than that of melanin production, melanin content per cell would decrease. Thus from the earlier findings (Hu et al., 1995) we are able to conclude that several factors control the complex melanogenesis process in biological systems and if we are capable of controlling these factors in culture, melanogenesis could be controlled in culture too. However, the chances are mutations are always higher in culture than the natural biological system.
To gain more insight into the changes of melanin composition during culture we investigated the influence of L-cystine, α-MSH and nitric oxide on Hanwoo muzzle melanocytes. It was observed that the total melanin and eumelanin production varies with treatments in different breeds (Table 1). Stimulation of melanogenesis by α-MSH was reported to favor a switch to the production of greater levels of eumelanogenic melanin and an increase in tyrosinase activity has been considered to be one of its mechanisms (Hunt et al., 1995; Thody and Graham, 1998). Besides α-SH, Nitric oxide (Romero-Graillet et al., 1998; Sasaki et al., 2000) is also a reported melanogenic factor and its also reported that nitric oxide contributes to eumelanogenesis enhancement. Our results are in complete agreement with studies of Romero-Graillet et al. (1998) and Sasaki et al. (2000) especially in case of Hanwoo brown melanocytes.
On the other hand an increase in total melanin content in Hanwoo brown and a significant (p<0.05) decrease of eumelanin content in Hanwoo black with L-cystine was observed (Table 1) in the current study. A decline in eumelanin with L-cystine has also been explained earlier (Hunt et al., 1995) and was demonstrated that L-cystine plays important role in pheomelanin production during melanogenesis. Moreover, mammalian pigmentation is controlled by the concerted action of TYR, TYRP1 and TYRP-2 in producing eumelanin and pheomelanin in melanocytes. We analyzed and found that Tyr is down regulated by L-cystine and up regulated by the nitric oxide and α-MSH in all the three breeds of Hawnoo. Tyrosinase is the rate-limiting enzyme in the melanogenesis pathway and high levels of this enzyme are required for the production of eumelanin, whereas low enzyme levels result in the production of pheomelanin (Hearing and Tsukamoto, 1991; Jackson, 1993). Tyrosinase activity is regulated by the melanocortin 1 receptor (MC1R or α-MSHR), whose stimulation by a-melanocyte-stimulating hormone leads to the production of eumelanin (Burchill et al., 1986).
Pheomelanin is produced either in absence of α-MSH stimulation or as result of a non-functional MC1R receptor (Robbins et al., 1993). Furthermore, eumelanogenesis is thought to be stimulated by the induction of tyrosinase-related enzymes (TRP-1 and TRP-2). It has also been reported that α-MSH has little effect on the expression of TRP-1 or TRP-2. Conversely, L-cystine concentration also has a strong influence on tyrosinase activity (Sieji et al., 1969). Seiji et al. (1969) demonstrated the direct inhibition of tyrosinase by cystine under in vitro condition. Whereas, the inactivation of the enzyme by the interaction of sulfhydryl compounds has been demonstrated by different authors (Jerjil et al., 1984; Jara et al., 1988; del et al., 1993). The regulation role of sulfhydryl compounds has already been described by Flesch et al (Flesch and Rothman, 1948). Del Marmol et al. (Del et al., 1993) have also shown that by the depletion of cystine in (hams F-10) medium of melanoma cell lines, tyrosinase activity was increased.
Interestingly, our findings are consistent with previous results on the levels of tyrosinase expression in all selected breeds. However, at present there is no adequate explanation as to why black and brindle cattle melanocytes failed to show any response to nitric oxide and α-MSH. It appeared to us that basal levels of tyrosinase and melanin content are unrelated to each other. Moreover, in the present study no convincing reason was found for the suppression of eumelanin production in black cattle by α-MSH. Additionally, death of melanocytes has been demonstrated earlier in culture (Nordlund et al., 2006) which might be due to accumulation of melanin that becomes toxic to the cells. Therefore, a probable reason for the death of melanocytes observed in the present study may be partly due to the bursting of cells in culture and partly by melanin toxicity. Actually it is already reported in the literature (Wick et al., 1977) that L-dopa, an intermediate in the biosynthesis of the pigment melanin, has highly selective inhibition of growth for pigmented cell lines as compared to the no-pigmented cells. There is a correlation between toxicity and the extent of incorporation of radioactively labeled L-dopa by each line. However, it is difficult to conclude from the present study that the real cause of cell toxicity is melanin accumulation. Further detailed investigations would be required.
In summary, the present investigation demonstrates that the nature of pigmentation is dependent on the stimulus. Eumelanogenesis increases significantly with α-MSH and nitric oxide which is accompanied by increase in tyrosinase activity. Finally, it can be concluded that melanogenesis can be stimulated in culture by the melanogenic factors viz., α-MSH, nitric oxide and L-cystine in muzzle melanocytes of Hanwoo cattle.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







