Methane Production of Different Forages in In vitro Ruminal Fermentation
Article information
Abstract
An in vitro rumen batch culture study was completed to compare effects of common grasses, leguminous shrubs and non-leguminous shrubs used for livestock grazing in Australia and Ghana on CH4 production and fermentation characteristics. Grass species included Andropodon gayanus, Brachiaria ruziziensis and Pennisetum purpureum. Leguminous shrub species included Cajanus cajan, Cratylia argentea, Gliricidia sepium, Leucaena leucocephala and Stylosanthes guianensis and non-leguminous shrub species included Annona senegalensis, Moringa oleifera, Securinega virosa and Vitellaria paradoxa. Leaves were harvested, dried at 55°C and ground through a 1 mm screen. Serum bottles containing 500 mg of forage, modified McDougall’s buffer and rumen fluid were incubated under anaerobic conditions at 39°C for 24 h. Samples of each forage type were removed after 0, 2, 6, 12 and 24 h of incubation for determination of cumulative gas production. Methane production, ammonia concentration and proportions of VFA were measured at 24 h. Concentration of aNDF (g/kg DM) ranged from 671 to 713 (grasses), 377 to 590 (leguminous shrubs) and 288 to 517 (non-leguminous shrubs). After 24 h of in vitro incubation, cumulative gas, CH4 production, ammonia concentration, proportion of propionate in VFA and IVDMD differed (p<0.05) within each forage type. B. ruziziensis and G. sepium produced the highest cumulative gas, IVDMD, total VFA, proportion of propionate in VFA and the lowest A:P ratios within their forage types. Consequently, these two species produced moderate CH4 emissions without compromising digestion. Grazing of these two species may be a strategy to reduce CH4 emissions however further assessment in in vivo trials and at different stages of maturity is recommended.
INTRODUCTION
Methane emissions from ruminant livestock have increased five fold over the last century (Johnson et al., 2000) and now constitute ~15% of global CH4 emissions (McAllister et al., 1996). In Australia, enteric CH4 emissions account for 10.5% of the national greenhouse gas emission (Grainger et al., 2008). Methane production also represents a loss of 2 to 12% of dietary energy in ruminants (Johnson and Johnson, 1995), this is especially the case in Australia with the consumption of high fibre pastures producing ~1 t CH4/t of liveweight gain (Leng, 1993). However, improving forage quality (i.e. increasing dietary starch content) through the supplementation of alternative forages, such as leguminous and non-leguminous shrubs, has the potential to reduce CH4 emissions per kg animal product as a result of increased diet digestibility and a shortened duration of feeding (Beauchemin et al., 2009). Dietary strategies such as this, have been successful in manipulating methanogenesis, at least in the short term, through either i) the direct inhibition of methanogens, ii) reducing the production of hydrogen in the rumen or iii) providing alternative sinks for the disposal of hydrogen (Beauchemin et al., 2009). For example, secondary plant compounds (e.g.: condensed tannins and saponins) have been shown to reduce enteric CH4 emissions through the direct inhibition of methanogens (Carulla et al., 2005). Similarly, the inclusion of high starch feedstuffs, favour the production of propionate and reduce ruminal pH, thus inhibiting methanogen and protozoal growth (Boadi et al., 2004).
The objective of this study was to characterise common grasses, leguminous shrubs and non-leguminous shrubs used for livestock grazing in parts of Australia and Ghana based on the methanogenic activity of ruminal bacteria (CH4 production at 24 h) using batch culture in vitro fermentation and to identify species which produce favourable fermentation profiles.
MATERIALS AND METHODS
Batch culture in vitro incubation
Twelve forage species were examined using batch culture in vitro fermentation. Grass species were Andropodon gayanus, Brachiaria ruziziensis and Pennisetum purpureum. Leguminous shrub species consisted of Cajanus cajan, Cratylia argentea, Gliricidia sepium, Leucaena leucocephala and Stylosanthes guianensis and non-leguminous shrub species were Annona senegalensis, Moringa oleifera, Securinega virosa and Vitellaria paradoxa. The forages were collected from native pastures near the Savanna Agricultural Research Institute at Nyankpala in the Guinea Savanna zone of Northern Ghana (9°54′N, 0°59′W). The location is characterized by a monomodal rainfall pattern with mean annual rainfall of 1,140 mm. Leaf samples of all leguminous and non-leguminous shrubs, and grasses were re-growths and collected ~60 d after the start of the rainy season. After harvest, leaves were dried at 55°C and ground through a 1 mm screen prior to in vitro incubation.
Rumen liquor was obtained from three ruminally fistulated lactating Holstein dairy cows (617±8.9 kg body weight; 45±13.1 d in milk) fed a total mixed ration containing 180 g crude protein (CP)/kg DM and 330 g neutral detergent fibre (aNDF)/kg DM. Cows were cared for in accordance with guidelines of the Canadian Council on Animal Care (1993). Ruminal liquor was collected from the anterior dorsal, anterior ventral, medium ventral, posterior dorsal and posterior ventral locations within the rumen 2 h after the morning feeding. Pooled ruminal fluid (e.g.: from the three cows) was strained through 4 layers of cheesecloth into an insulated thermos and immediately transported to the laboratory. Strained ruminal fluid was continuously purged under O2-free N gas and kept at 39°C in a water bath prior to use in in vitro incubations. The liquor was used for inoculum of in vitro bottles within 20 min of collection.
In vitro incubations were in 50 ml bottles fitted with rubber stoppers to prevent escape of fermentation gases (Wang et al., 2000). Bottles were placed in an incubator (Forma Scientific, model 39419-1, Marietta, OH, USA) with a capacity for 204 bottles. Incubator temperature was 39±0.5°C and bottles were affixed to a rotary shaker platform at 120 oscillations/min (Lab-Line Instruments Inc, Melrose Park, IL, USA). On the day before incubation, 0.5 g DM of each forage sample (n = 2) was weighed into each bottle (n = 3 for each treatment). On the day of incubation, bottles containing substrate were warmed to 39°C in the incubator for 60 min, and gassed with N2 before adding 20 ml of a 3:1 mixture of buffer:rumen fluid saturated with CO2 with 0.5 ml cysteine sulphide reducing agent (Wang et al., 2000). Bottles were capped and placed in the incubator at 39°C. Three bottles containing inoculum only were also included at each time point as a blank control.
Gas samples were measured at 2, 6, 12 and 24 h of incubation. The procedure for sampling bottles was: 39 bottles (3 replicates×12 treatments+3 blanks) were removed from the incubator and gas production from each bottle was immediately determined using a water displacement apparatus (Fedorak and Hrudey, 1983). For CH4 determination, 20 ml of headspace gas was collected from the bottles at 24 h by inserting a 20 ml syringe through the septum and immediately transferring it into a 6.8 ml evacuated exetainer (Labco Ltd., High Wycombe, England) thus ensuring a positive pressure in the containers. A 3 ml subsample of gas was removed from each exetainer and analyzed for CH4 by gas chromatography as described by Chaves et al. (2006). Methane was expressed as mg CH4/g digested DM and total net gas production in ml/g DM incubated (Chaves et al., 2006).
At 24 h of incubation, incubation bottles (n = 3) of each forage were withdrawn from the incubator and put in ice to stop fermentation, and then processed for determination of volatile fatty acids (VFA), ammonia and in vitro DM disappearance (IVDMD) as described by Wang et al. (2008).
Determination ammonia
A sub-sample (1.5 ml) of the media culture from each bottle were transferred to 2 ml micro-centrifuge tubes containing 150 μl trichloroacetic acid (TCA; 0.25%; w/v) and centrifuge at 14,000×g for ten minutes (Spectrafuse 16M, National Labnet Co., Edison, NJ, USA) to precipitate particulate matter. The supernatant was transferred into 2 ml microcentrifuge tubes and stored frozen at −20°C until it was analyzed for ammonia-N by phenyl-hypochlorite reaction (Weatherburn, 1967). Blank 0 h samples were also used to analyze for ammonia and to calculate the net ammonia-N production.
Determination of volatile fatty acids (VFA)
An additional sub-sample (1.5 ml) of the culture in each bottle was also removed and 150 μl of metaphosphoric acid (20%; w/v) was added before the samples were centrifuged in the same manner as the ammonia analysis. The supernatant was stored frozen at −20°C and later analyse for VFA concentrations by high-performance liquid chromatography (HPLC; SHIMADZU model SPD-10A VP with an Ultra UV Detector waves lengths of 210 nm) equipped with a SCR-101 H silica 30 cm×7.9 mm×1 μm capillary column. Blank 0 h samples were also analyzed for VFA and to determine the net total VFA production.
In vitro DM disappearance
Following the collection of the supernatants samples for ammonia and VFA analyses after 24 h of incubation, the contents of the incubation bottles were transferred into preweighed 50 ml centrifuge tubes, rinsed with distilled water into the tubes, and centrifuged twice at 500×g for 10 min at 4°C. The supernatant was discarded, and the precipitate dried at 55°C for 48 h and weighed to estimate the in vitro DM disappearance (IVDMD).
Chemical composition
Duplicate samples of each forage were analysed for neutral detergent fibre (aNDF) with a heat stable α-amylase and sodium sulphite (Mertens, 2002) and for acid detergent fibre (ADF) using method 973.18 (AOAC, 1990). Concentrations of aNDF and ADF are expressed inclusive of residual ash. Lignin (sa) was completed on ADF residues by solubilisation with sulphuric acid (Robertson and Van Soest, 1981). Ash was determined after 2 h of oxidation at 600°C in a muffle furnace using method 942 (AOAC, 1990). Samples were re-ground using a ball grinder (Mixer Mill MM2000, Retsch, Haan, Germany) for determination of N by flash combustion with gas chromatography and thermal conductivity detection (Carlo Erba Instruments, Milan, Italy).
Statistical analyses and calculation
Methane production was calculated as:
according to Chaves et al. (2006).
In vitro data were analyzed as a completely randomized design using the MIXED procedure of SAS (2011). Means within forage family (e.g., leguminous shrubs, non-leguminous shrubs or grasses) were compared using the least squares mean linear hypothesis test included as fixed terms. Results are reported as least squares means (±SEM). Significance was declared at p≤0.05.
RESULTS AND DISCUSSION
Concentrations of crude protein (CP; g kg−1 DM) and NDF (g kg−1 DM) in P. purpureum, G. sepium, and M. oleifera are comparable to previously reported values of 96 and 709 (Nogueira Filho et al., 2000); 92 and 421 (Abdulrazak et al., 1997) and 214 and 333 (Njidda and Nasiru, 2010), respectively (Table 1). Dietary crude protein and NDF concentrations influence the amount of substrate organic matter (OM) fermented and the short chain fatty acids that are produced (Njidda and Nasiru, 2010). Crude protein concentrations below the threshold of 70 g CP kg−1 DM, restrict microbial activity due to a lack of nitrogen (Hariadi and Santoso, 2010). However, protein levels above this threshold, as seen in this study, are considered to enhance microbial multiplication in the rumen, thus improving fermentation (Njidda and Nasiru, 2010). Similarly, ammonia accumulation varied (p<0.05) within legume and shrub species, yet remained above the threshold value for both maximum microbial growth (Satter and Slyter, 1974) and fibre digestion (Hariadi and Santoso, 2010). This suggests that CP concentrations did not impede gas production or forage digestibility.
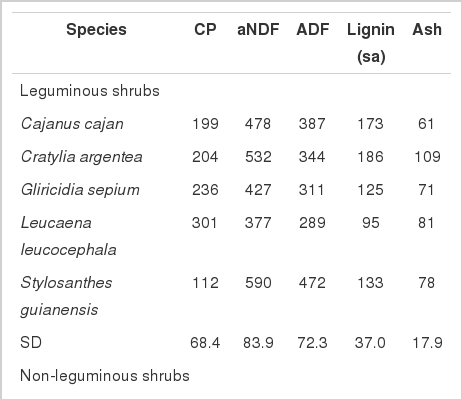
Chemical composition of forage species expressed in g/kg DM) used in the study
In vitro DMD and cumulative gas production (ml/g DM) also differed (p<0.05) within forage type in the current study. B. ruziziensis, G. sepium and M. oleifera produced the highest (p<0.05) cumulative gas and had the highest IVDMD within the grasses, leguminous and non-leguminous shrubs, respectively (Table 2) indicating that these species are highly fermentable. It is widely accepted that feedstuffs which have higher gas production and IVDMD tend to have higher CH4 production per gram DM incubated (Durmic et al., 2010; Njidda and Nasiru, 2010; Jayanegara et al., 2011). This trend was evident in non-leguminous shrub species, in addition to P. purpureum and S. guianensis (data not presented). This relationship is suggested to be a reflection of feed digestibility (Demeyer and Van Nevel, 1975; Holtshausen et al., 2009), in particular NDF concentration (Doane et al., 1997). Increasing cell wall content (i.e. NDF and ADF) is considered to suppress microbial activity through a reduction in the availability of rapidly fermented carbohydrates (Wilson and Hatfield, 1997). This is turn, is negatively correlated with gas production (Njidda and Nasiru, 2010). Similarly, in in vivo trials increasing fibre content is considered to reduce forage intake, ruminal residence time (Beauchemin et al., 2009; Archimède et al., 2011) and fermentability (Wilson and Mertens, 1995) resulting in an increase in CH4 production per unit of OM digested (McAllister et al., 1996; Valenciaga et al., 2009).

Cumulative gas and methane production, ammonia accumulation, concentration and proportions of volatile fatty acids (VFA) and in vitro dry matter (DM) disappearance (IVDMD) of 24 h in vitro ruminal fermentation of different forage species
In the current study, several forage species were able to favourably modulate rumen fermentation to produce CH4 concentrations lower than expected when considering their cumulative gas production and IVDMD. The most notable of which, G. sepium and B. ruziziensis, produced moderate amounts of CH4 compared with other species in their forage type, despite having the highest cumulative gas production, IVDMD and total VFA production. A strong positive relationship was observed between IVDMD and total VFA production, with total VFA production the greatest in species with high IVDMD, as would be expected. Interestingly, both G. sepium and B. ruziziensis, produced the highest proportion of propionate, and lowest acetate:propionate ratios, within their forage types.
Generally, the type of carbohydrate present in the forage is thought to dictate CH4 production via shifts in the ruminal microbial population (Johnson and Johnson, 1995). High soluble carbohydrate content is suggested to promote the production of propionate in the rumen, lower ruminal pH and inhibit methanogen growth thereby reducing CH4 production per unit of OM fermented (Van Kessel and Russell, 1996). Boadi and Wittenberg (2002) found CH4 production (L CH4/kg digestible OM intake) was 25% higher for low versus medium or high nutritional quality diets in a feeding study where intake was restricted to 2% of body weight. Lower CH4 losses with the high quality diet were expected as the lower fibre content shifted fermentation towards production of propionate. Chaves et al. (2006) demonstrated that diet quality affected CH4 production in an in vitro study where low concentrations of non-fibrous carbohydrates in both legumes (e.g., alfalfa) and grasses contributed to a low IVDMD and consequently high CH4 production per gram digested DM. Similarly, Boadi et al. (2002) showed that steers grazing early season pasture produced up to 45% less CH4 than those grazing in the mid and late season. However, in the current study, no correlation between NDF concentration and CH4 production was observed.
The presence of plant secondary metabolites (e.g. condensed tannins and saponins) may account for this, although, their presence was not examined in the current study. Tannins, common in tropical legumes (Waghorn, 2008), have been shown to reduce CH4 production by reducing fibre digestion (Tiemann et al., 2008); binding with proteins thus reducing degradation of the plant protein in the rumen and through the direct inhibition of the growth of methanogens (Tavendale et al., 2005). A study by Pinares-Patino et al. (2003) reported a reduction in CH4 emissions from grazing sheep, from 8% GEI to 3% GEI, following consumption of the tannin rich forage, Lotus corniculatus. Similarly, Carulla et al. (2005) found that an extract of Acacia mearnsii (61.5% CT) reduced CH4 emissions by 12% without decreasing fibre digestibility.
G. sepium and B. ruziziensis, were able to successfully modulate rumen fermentation characteristics, such that moderate CH4 emissions were produced without compromising forage digestion. Consequently, grazing of these two species may be a strategy to reduce CH4 emissions in Australia. However, as our forages were examined at only one stage of maturity, any effects of NDF concentration on CH4 production were not realised, as such there is a need for further assessment of fermentation characteristics of these species at various stages of maturity. Similarly, the relative scarcity of data on CH4 production from in vivo trials, where ruminants are fed tropical forages, suggests a need for additional studies.