




INTRODUCTION
In vertebrates, most biochemical, physiological and behavioural parameters exhibit daily fluctuations. These circadian rhythms are generated by endogenous oscillators, termed the ‚Äėcircadian clock‚Äô with a period very close to 24 hours, and are internally generated to anticipate environmental changes associated with solar days [1]. In vertebrates, the pineal gland is a neuroendocrine organ that converts environmental photoperiodic information into biochemical messenger encoding darkness‚ÄĒmelatonin (N-acetyl-5-methoxytryptamine). The day/night rhythm of melatonin secretion reliably reflects environmental light conditions and is independent from the behaviour. Changes in night length, caused by natural succession of seasons, are translated into fluctuation in the duration and/or amplitude of the melatonin surge [2]. A conservative feature of vertebrates is that the plasma level of melatonin increases at night, whereas diurnal levels of this hormone are relatively low [3]. In all studied mammalian species, melatonin secretion is stimulated by norepinephrine (NE), which is released from sympathetic nerve fibres exclusively at night [2]. Biosynthesis of melatonin in pinealocytes is a multistep sequence of reactions that starts with the hydroxylation of the amino acid tryptophan to 5-hydroxytryptophan (5-HTP) by the tryptophan hydroxylase (TPH). Next, aromatic amino acid decarboxylase (DDC) converts 5-HTP to 5-hydroxytryptamine (5-HT; serotonin). Then, 5-HT is transformed to N-acetylserotonin by arylalkylamine-N-acetyltransferase (AANAT). Finally, N-acetylserotonin is converted to melatonin by the hydroxyindole-O-methyltransferase (HIOMT) [4]. Circulating levels of melatonin in the bloodstream fully reflect pineal secretory activity because melatonin is not stored within pinealocytes, but it freely diffuses out of the cells into blood capillaries immediately after its formation [2].
It is well established that amongst the numerous actions of melatonin, it plays the role of an immune modulator regulating the development, differentiation and function of lymphoid tissues. Moreover, diurnal and seasonal changes in immune function are thought to directly reflect changes in pineal melatonin production, which suggests an important role of circulating melatonin in the development and maintenance of immune system [5]. The pineal gland is likely to participate in the innate immune response because it expresses mRNA encoding transcripts for all ten members of the toll-like receptor (TLR) family. Therefore, the function of this gland may be affected by a number of pathogen-associated molecular patterns recognised by these receptors [6]. Interactions occurring between the immune system and the pineal gland appear to be bidirectional. However, the feedback action of the activated immune system on melatonin biosynthesis in the pineal gland is still poorly understood.
This study was designed to determine the effects of an immune/inflammatory challenge induced by intravenous (i.v.) injection of bacterial endotoxin‚ÄĒlipopolysaccharide (LPS)‚ÄĒon melatonin release and the expression of the enzymes of the melatonin biosynthetic pathway in sheep during two distinct photoperiodic conditions: a short-night (SN: long-day, 8:16) and long-night (LN: short-day, 16:8) period. Moreover, the influence of LPS treatment on the gene expression of proinflammatory cytokines and their corresponding receptors in the pineal gland was also studied.
MATERIAL AND METHODS
Animals and experimental design
The study was performed on adult, three-year-old blackface ewes (n = 24) during two different photoperiods: in May/June during a SN (night:day, 8:16) period and in November/December during a LN (night:day, 16:8) period. Experiments started one month before the summer and winter solstices, respectively. Animals were maintained indoors in individual pens in controlled temperature (16¬įC to 18¬įC) and humidity (60% to 70%). The ewes were exposed to the natural daylight present at 52¬įN latitude and 21¬įE longitude. The ewes were in good condition and were kept under veterinary care. The animals were acclimated to the experimental conditions for one month. Ewes were always within visual contact with other members of the flock to prevent isolation stress. The animals were fed a consistent diet of commercial concentrates with hay and water available ad libitum. All animals had venous catheters implanted into their jugular vein the day before the experiment. All animal procedures were approved by the Local Ethics Committee of Warsaw University of Life Sciences‚ÄĒSGGW (Warsaw, Poland; authorisation no. 59/2011).
In both experiments, animals (n = 12) were randomly divided into two groups: control (n = 6) and LPS-treated (n = 6) one. The experimental procedures were performed at night, in the darkness and in the presence of a red light. Two hours after sunset, animals received an i.v. injection of LPS from Escherichia coli 055:B5 purified by phenol extraction, characterized by <3% impurity with protein (cat no. L2880; Sigma-Aldrich, St. Louis, MO, USA) dissolved in saline (0.9% w/v NaCl) (Baxter, Deerfield, IL, USA) at a dose of 400 ng/kg. The control animals received an i.v. injection of saline (2.5 mL). Blood samples were collected at 15-min intervals, starting one hour after sunset and continuing for 3 hours after the treatment. Blood samples were collected into heparinised tubes and immediately centrifuged in an MPW 260RH centrifuge (MPW Med. Instruments, Warsaw, Poland) for 10 min at 1,000√óg at 4¬įC. Plasma was stored at ‚ąí80¬įC until assayed. The ewes were euthanized 3 hours after LPS/NaCl injection at 3 am during the SN period and 10 pm. during the LN period. The brains were immediately removed from the skulls, and the pineal gland was dissected into three parts, frozen in liquid nitrogen and stored at ‚ąí80¬įC until assay.
Radioimmunoassay of plasma hormones
Melatonin was assayed in blood plasma according to the method of Fraser et al [7] and as modified in our laboratory using ovine anti-melatonin serum (Dr A. Foldes, CSRIO, Canberra, Australia). Synthetic melatonin (Sigma-Aldrich, USA) was used as a standard and [O-methyl-3H]-melatonin (Amersham PLC, Amersham, UK) as a tracer. The sensitivity of the assay was 17¬Ī8 pg/mL, and the intra- and interassay coefficients of variation were 11% and 13%, respectively.
Cortisol concentrations in the plasma were determined via radioimmunoassay according to Kokot and Stupnicki [8] using rabbit anti-cortisol antisera (R/75) and an high-performance liquid chromatography (HPLC)-grade cortisol standard (Sigma-Aldrich, USA). The assay sensitivity was 1 ng/mL, and the intra- and interassay coefficients of variation for cortisol were 9% and 12%, respectively.
Determination of serotonin (5-HT) and its metabolite 5-hydroxyl-indole-3-acetic acid content in the pineal gland
Pineal glands were analysed for 5-HT and 5-hydroxyl-indole-3-acetic acid (5-HIAA) contents using HPLC with electrochemical detection. Final concentrations were calculated relative to the recovery of the internal standard 5-hydroxyindole (Sigma-Aldrich, USA) and expressed as pg/mg of tissue. The analytical procedure was as followed. Briefly, pineal glands were thawed, homogenised at 0¬įC in 0.1 M HClO4 (1:10 w/V) (Sigma-Aldrich, USA) with 200 ng of 5-hydroxyindole added as an internal standard, and samples were centrifuged in a SIGMA 1‚Äď14K centrifuge (Sigma Laborzentrifugen GmbH, Osterode am Harz, Germany) at 12,000 √óg for 15 min. The concentrations of 5-HT and 5-HIAA were determined via HPLC using a Waters 515 system (Waters Corporation, Milford, MA, USA) coupled to an electrochemical detector (Hewlett Packard 1049A, HP Inc., Palo Alto, CA, USA) equipped with a glassy carbon working electrode and an Ag/AgCl reference electrode. The electrochemical detector was set at an oxidative potential of 0.650 V. The supernatants (50 őľL) were injected into an LC-18-DB (15 cm√ó4.6 mm ID, 5 őľm) Supelco column (Sigma-Aldrich, USA) protected by a SUPELCOSIL LC-18-DB Supelguard 2-cm pre-column (5 őľm particle size; Sigma-Aldrich, USA) and were eluded isocratically with a mobile phase consisting of 0.01 mol/L NaCl (Sigma-Aldrich, USA), 0.001 mol/L ethylenediaminetetraacetic acid (Sigma-Aldrich, USA) and 12% CH3OH (Sigma-Aldrich, USA). The pH of the mobile phase was 3.6, and the flow rate was set at 0.8 mL/min. The limit of detection was 10 pg/50 őľL for 5-HT and 5 pg/50 őľL for 5-HIAA.
Determination of intracellular concentrations of cAMP in the pineal gland
Concentrations of cAMP were determined in pineal tissue using an RIA kit from Immunotech (Beckman Coulter, Brea, CA, USA) according to the manufacturer‚Äôs protocol. The applied assay had an analytical sensitivity of 0.2 nM, an intra-assay coefficient of variation (CV) of 11% and an inter-assay CV of 16%. Pineal tissues were homogenised in 300 őľL of 0.01 M chilled phosphate-buffered saline (Sigma-Aldrich, USA) using a TissueLyser LT (Qiagen, Hilden, Germany). Next, the homogenates were centrifuged at 6,000√óg at 4¬įC for 5 min, and the supernatants were collected. The protein concentrations of the samples were quantified using the Pierce Coomassie Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer‚Äôs protocol. The measurement of the absorbance at 595 nm was performed using a VersaMax reader (Molecular Devices LLC., Sunnyvale, CA, USA). Samples were diluted (1:1) with assay diluent anPd added to antibody coated tubes, which were then incubated overnight at +4¬įC with I125-labelled cAMP tracer. Radioactivity was measured using a Packard Cobra II Gamma Counter (Packard Instrument Company, Inc., Meriden, CT, USA). A standard curve was constructed using a log-linear curve fit with B/Bo (%) (y-axis, where B is the average cpm of the paired standards and Bo is the cpm of total activity) against cAMP concentration (x-axis). Values were normalised to the amount of protein (mg) present in the sample.
Isolation of mRNA and protein from pineal glands
Total RNA and protein from explants were isolated using a NucleoSpin RNA/Protein Kit (MACHEREY-NAGEL Gmbh & Co., D√ľren, Germany). All isolation steps were performed according to the manufacturer‚Äôs protocols. The purity and concentration of the RNA extractions were quantified by measuring the optical density at 230, 260, and 280 nm in a NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific Inc., USA). RNA integrity was confirmed by electrophoresis using a 1% agarose gel (Reducta NU, PRONA Marine Research Institute, Vigo, Spain) stained with ethidium bromide (Sigma-Aldrich, USA).
Real-time polymerase chain reaction assay
To synthesise cDNA, the Maxima First Strand cDNA Synthesis Kit for quantitative reverse transcription polymerase chain reaction (RT-qPCR) (Thermo Fisher Scientific, USA) and 2 őľg of total RNA were used. Real-time qRT-PCR was performed using the HOT FIREPol EvaGreen qPCR Mix Plus (Solis BioDyne, Tartu, Estonia) and HPLC-grade oligonucleotide primers (Genomed, Warsaw, Poland). Primer sequences were designed using Primer3 software (http://primer3.ut.ee) (Table 1). Each reaction mixture (total volume: 20 őľL) contained 4 őľL of PCR Master Mix (5√ó), 14 őľL of RNase-free water, 1 őľL of primers (0.5 őľL each primer, working concentration 0.25 őľM) and 1 őľL of the cDNA template. Reactions were run on a Rotor-Gene Q instrument (Qiagen, Germany), and the following protocol was used: 95¬įC for 15 min and 30 cycles of denaturation at 95¬įC for 10 s, annealing at 60¬įC for 20 s, and extension at 72¬įC for 10 s. A final melting curve analysis was performed to confirm the specificity of amplification.
Relative gene expression was calculated using the comparative quantification option [9] of the Rotor Gene Q software version 1.7 (Qiagen, Germany). Three housekeeping genes were examined: glyceraldehyde-3-phosphate dehydrogenase, ő≤-actin (ACTB), and histone deacetylase 1. The mean expression of these three housekeeping genes was used to normalise the expression of the analysed genes. The results are presented in arbitrary units as the ratio of target gene expression to the mean expression of the housekeeping genes.
Western-Blot assay
Before electrophoresis, the protein concentrations of samples isolated using the NucleoSpin RNA/Protein Kit (MACHEREY-NAGEL Gmbh & Co., Germany) were quantified using a Protein Quantification Assay Kit (MACHEREY-NAGEL Gmbh & Co., Germany). The appropriate volume of molecular grade water (Sigma-Aldrich, USA) was added to a volume of sample containing 50 őľg of total protein to bring the total sample volume to 20 őľL. Next, 19 őľL of Laemmli buffer (Sigma-Aldrich, USA) and 1 őľL of ő≤‚Äďmercaptoethanol (Sigma-Aldrich, USA) were added. Such mixtures were boiled for 3 min. Electrophoresis was then performed in the presence of molecular weight markers (Spectra Multicolor Broad Range Protein Ladder, Thermo Fisher Scientific Inc., USA). Denatured samples and molecular weight standards were loaded onto 4% to 12% polyacrylamide gels and subjected to electrophoresis in a Tris-glycine running buffer using the Protean II xi Cell (Bio-Rad Laboratories, Inc., Hercules, CA, USA) according to the manufacturer‚Äôs instructions. Next, proteins were transferred in Tris-glycine blotting buffer to polyvinylidene difluoride membranes (Immobilon-P [0.45 őľm], Merck KGaA, Darmstadt, Germany) using the Trans-Blot SD Semi-Dry Transfer Cell (Bio-Rad Laboratories, Inc., USA) for 30 min at 20 V. The membranes were blocked for 1 h at room temperature in blocking buffer made up of Tris buffered saline at pH 7.5 with 0.05% Tween-20 (TBST) (Sigma-Aldrich, USA) containing 3% bovine serum albumin fraction V (Sigma-Aldrich, USA). Next, membranes were incubated overnight at 4¬įC with the following primary antibodies: goat anti-TPH polyclonal antibody (cat no. sc-15116, Santa Cruz Biotechnology Inc., Dallas, TX, USA), rabbit anti-DDC polyclonal antibody (cat no. sc-99203, Santa Cruz Biotechnology Inc., USA), goat anti-AANAT polyclonal antibody (cat no. sc-55612, Santa Cruz Biotechnology Inc., USA), rabbit anti-HIOMT polyclonal antibody (cat no. bs-6961R, Bioss Inc., Woburn, MA, USA), and mouse anti-ACTB monoclonal antibody (cat no. sc-47778, Santa Cruz Biotechnology Inc., USA) dissolved in blocking buffer at dilutions of 1:100, 1:200, 1:100, 1:200, and 1:1,000, respectively. After washing three times, membranes were incubated with the following secondary horseradish peroxidase (HRP) conjugated antibodies: bovine anti-rabbit immunoglobulin G (IgG)-HRP (cat no. sc-2379, Santa Cruz Biotechnology Inc., USA), donkey anti-goat IgG-HRP (cat no. sc-2304, Santa Cruz Biotechnology Inc., USA) and goat anti-mouse IgG1 heavy chain (HRP) (cat no. ab97240, Abcam, Cambridge, UK) dissolved in blocking buffer at a dilution of 1:10,000. After washing three times, the membranes were visualised using chromogenic detection with a Pierce 1-step 3,3‚Ä≤,5,5‚Ä≤-Tetramethylbenzidine blotting substrate solution (Thermo Fisher Scientific, USA). After visualisation, the membranes were dried and scanned using an EPSON Perfection V370 Photo scanner (Seiko Epson Corporation, Suwa, Japan). Densitometric analysis of the scanned membrane was performed using the software ImageJ (Research Services Branch, National Institute of Mental Health, Bethesda, MD, USA).
Statistical analysis
Statistical analyses were performed using the software STATISTICA 10 (StatSoft Inc., Tulsa, OK, USA). The raw data of the melatonin and cortisol concentration in blood, after verification of the assumptions of normality (Shapiro-Wilk‚Äôs test), homogeneity of variances (Levene‚Äôs test), and sphericity (Mauchly‚Äôs test) were subjected to 2 (LPS or saline treated) by 2 (photoperiod: SN, LN) by 4 (sample collection time: 1 h, 2 h, 3 h, and 4 h) repeated measures analysis of variance (ANOVA) followed by Fisher‚Äôs least significant difference post-hoc test. The sample collection time factor was treated as repeated measures factor on the same unit (within-subject). It should be noted that for melatonin and cortisol data, to identify treatment effects, the mean values for 1-hour periods were established and included in statistical analyses. On the other hand, the raw data on the concentration of 5-HT and 5-HIAA as well as the expression of genes and proteins, after passing normality and homogeneity of variances tests, were subjected to a two-way ANOVA to determine any significant influences of the two parameters (photoperiod and LPS-treatment), followed by Fisher‚Äôs least significant difference post-hoc test. The results are presented as the mean¬Īstandard error of the mean. Statistical significance was set at p<0.05.
RESULTS
Influence of LPS injection on melatonin and cortisol release
It was found that LPS-treated animals from LN photoperiods were characterised by lower (p<0.05) levels of circulating melatonin (162¬Ī11 pg/mL) compared with control animals from the LN (218¬Ī12 pg/mL) photoperiod during the first hour after treatment. However, no differences in the plasma levels of melatonin were found after this period in ewes tested during the SN photoperiod. On the other hand, endotoxin treatment reduced (p<0.05) the mean melatonin secretion in ewes during the SN photoperiod (108¬Ī4 pg/mL) only in the third hour after injection compared to control ewes (171¬Ī12 pg/mL)(Figure 1).
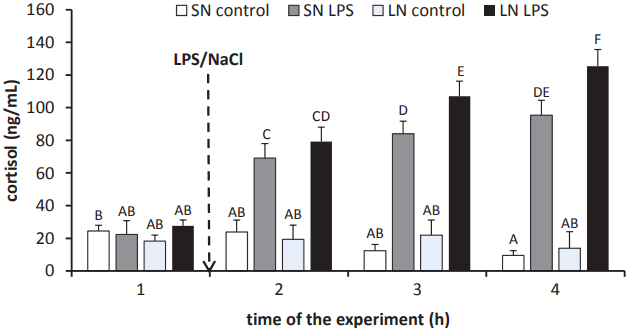
Treatment with LPS stimulated (p<0.05) cortisol release in ewes from both the SN and LN photoperiods starting from the first hour after treatment. Moreover, it was found that ewes in the LN photoperiod were characterised by higher (p<0.05) circulating concentrations of cortisol (107¬Ī9 ng/mL and 125¬Ī10 ng/mL) from the second hour on after LPS injection compared with levels in treated ewes from the SN photoperiod (84¬Ī8 ng/mL and 85¬Ī9 ng/mL, respectively). It should be noted that when in the LPS-treated animals from SN photoperiod the circulating level of cortisol was equal at the 2nd and 3rd h after the endotoxin injection, in the ewes from LN photoperiod blood concentration of cortisol was growing with each subsequent hour after LPS administration (Figure 2).
Effect of LPS injection on concentration of 5-HT and 5-HIAA in the pineal gland
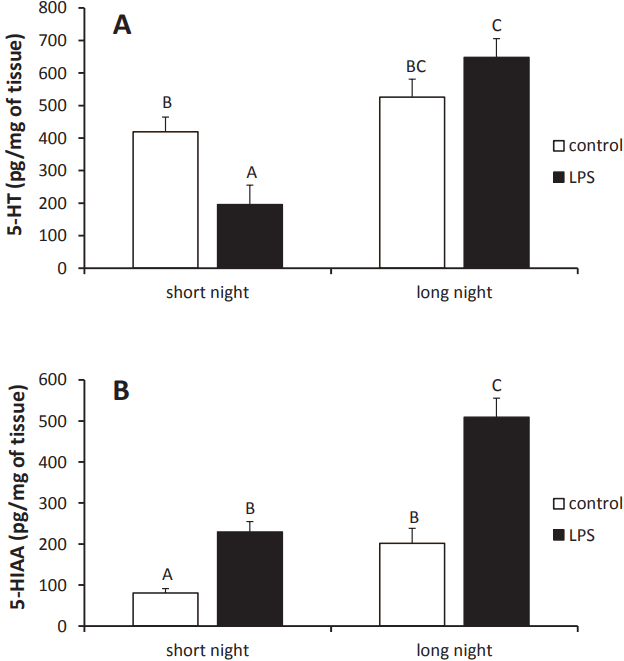
Injection of LPS reduced (p<0.05) 5-HT concentrations in the pineal tissue of animals from the SN (196¬Ī59 pg/mg of tissue) photoperiod compared with levels in control ewes (419¬Ī45 pg/mg of tissue). However, no effect of the LPS treatment on pineal 5-HT concentration was found in ewes from the LN photoperiod (Figure 3A).
The concentrations of 5-HIAA in the pineal glands of the control ewes were lower (p<0.05) during the SN (80¬Ī11 pg/mg of tissue) photoperiod than during the LN (201¬Ī36 pg/mg of tissue) photoperiod. Administration of LPS elevated (p<0.05) the concentration of 5-HIAA in the pineal glands of ewes during both the SN (229¬Ī25 pg/mg of tissue) and LN (509¬Ī46 pg/mg of tissue) photoperiods (Figure 3B).
Effect of LPS injection on concentration of cAMP in the pineal gland
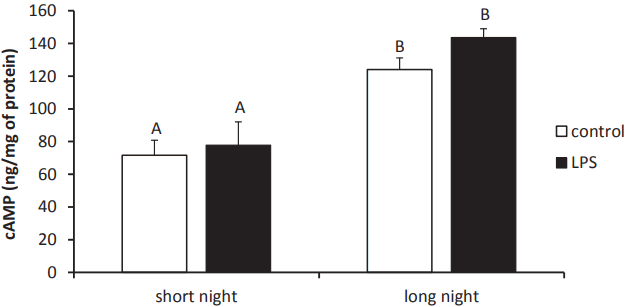
The concentrations of cAMP in the pineal glands collected from ewes during the SN (72¬Ī9 pg/mg of protein) photoperiod were lower (p<0.05) than from animals during the LN (124¬Ī7 pg/mg of protein) photoperiod. However, no effect of LPS treatment on the pineal cAMP content was found (Figure 4).
Influence of LPS on relative gene and protein expression of enzymes of the melatonin biosynthetic pathway
Peripheral injection of LPS decreased (p<0.05) the expression of TPH1 mRNA only in pineal tissues collected from ewes during the SN photoperiod but suppressed (p<0.05) pineal protein expression of the TPH regardless of photoperiod. Moreover, it was found that TPH protein expression was higher (p<0.05) during the LN compared to the SN photoperiod (Figure 5; panel A). Although LPS did not influence DDC expression both at gene and protein level, it was found that DDC was more strongly (p <0.05) expressed both at the protein and mRNA level in the pineal gland of ewes during LN compared to SN photoperiods (Figure 5; panel B). Endotoxin-induced inflammation lowered (p<0.05) the gene and protein expression of AANAT only in ewes from the SN photoperiod (Figure 5; panel C). No influence of LPS on HIOMT protein and gene expression was found for either photoperiod (Figure 5; panel D).
Effect of LPS administration on gene expression of toll-like receptor 4 and proinflammatory cytokine receptors in the pineal gland
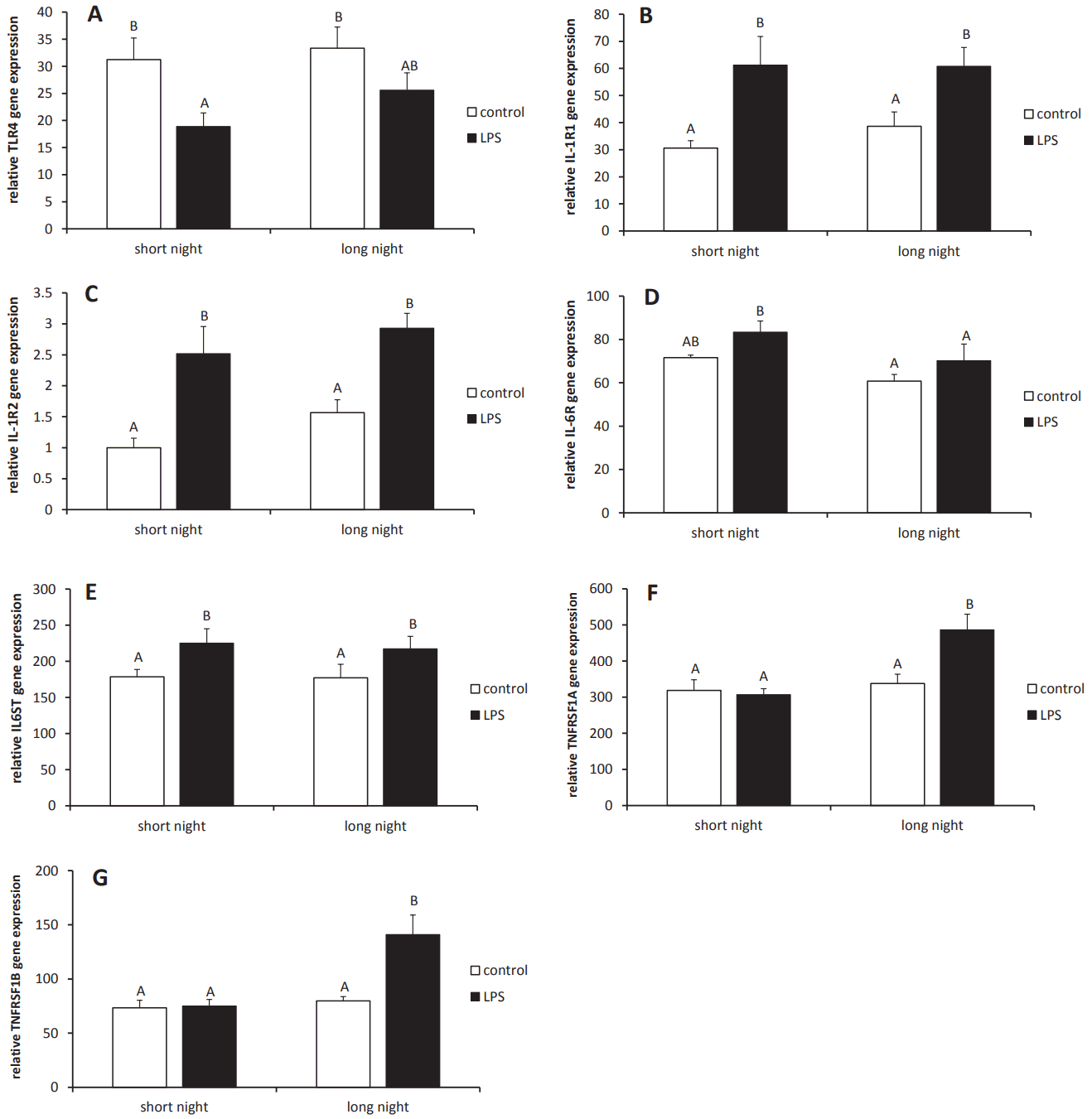
Intravenous administration of LPS suppressed (p<0.05) the expression of the toll-like receptor 4 (TLR4) gene in the pineal gland tissues collected from ewes during the SN photoperiod. However, regardless of the photoperiod, endotoxin treatment stimulated (p<0.05) expression of the genes encoding interleukin (IL) ‚ąí1 receptor 1 (IL1R1), IL1R2 and glycoprotein 130 (IL6ST), which are necessary for the transduction of IL-6R signals. Injection of LPS increased (p<0.05) mRNA expression for tumor necrosis factor (TNF) type 1 receptor (TNFRSF1A) and type 2 (TNFRSF1B) in the pineal glands from ewes during the LN photoperiod. No influence of endotoxin treatment was found on IL6R mRNA expression in the pineal gland (Figure 6).
DISCUSSION
It is the first study describing the effects of inflammation induced by a bacterial endotoxin on the nocturnal secretion of melatonin in ewes. The obtained results show that an immune/inflammatory challenge may affect the secretion of melatonin but this influence is dependent upon photoperiod. The administration of endotoxin decreased circulating levels of melatonin in ewes during the LN photoperiod, but this effect was limited to only the first hour after treatment. However, during the SN photoperiod, injection of LPS reduced melatonin secretion only in the third hour after treatment, and this effect was maintained until the end of the experiment. This suggests that the mechanisms affecting the nocturnal secretion of melatonin via inflammation are different during SN and LN photoperiods. The most important factor influencing immune-pineal interaction may be melatonin itself. Melatonin regulates inflammatory and immune processes and acts as both an activator and inhibitor of these responses. It is postulated that melatonin is an immune buffer, acting as a stimulant under basal or immunosuppressive conditions, or as an anti-inflammatory compound in the presence of exacerbated immune responses, such as acute inflammation [10]. Moreover, the effects of melatonin on the immune response may be dependent on the phase of this response. Melatonin appears to promote early phases of inflammation on the one hand and contributes to its attenuation on the other, thus avoiding complications of chronic inflammation [11]. This suggests that a high level of circulating melatonin could accelerate the course of the inflammatory response in ewes during the LN photoperiod, and therefore LPS-dependent suppression of melatonin secretion occurred quickly, just after the treatment. However, high concentrations of melatonin attenuate the immune response, which resulted in diminished inhibitory action on melatonin synthesis in the pineal gland. Otherwise, lower concentrations of circulating melatonin in ewes during the SN photoperiod might not affect the inflammatory response to such an extent as in the LN photoperiod. Therefore, the inhibitory influence of inflammation on melatonin secretion was observed later.
Endotoxin-induced inflammation may disturb secretory activity via different mechanisms involving diverse mediators including proinflammatory cytokines. The present study documented the expression of mRNAs encoding receptors for IL-1, IL-6, and TNFőĪ in the pineal gland. This suggests that these proinflammatory cytokines may affect the secretory activity of this gland. This is supported by our previous ex vivo study on ovine pineal glands explants [12] and in vivo study on sheep conducted in two different photoperiodic conditions [13] showed that IL-1ő≤ is the potent down-regulator of melatonin secretion. The lack of seasonal differences in the gene expression of proinflammatory cytokine receptors in the pineal gland under basal conditions suggests that the photoperiod does not influence the sensitivity of the pineal gland to the action of inflammatory mediators corresponding to these receptors. However, results indicating that expression of TNFRSF1A and TNFRSF1B was enhanced in the LPS-treated group suggest that, under inflammatory conditions, pineal glands are more sensitive to the action of TNFőĪ during LN than SN photoperiods. TNFőĪ inhibits AANAT mRNA expression as well as the synthesis of the melatonin precursor N-acetylserotonin [14]. The major pool of proinflammatory cytokines affecting the activity of melatonin secretion are circulating molecules reaching the pineal gland via peripheral blood and that a significant amount of these mediators could be synthesised directly in pineal tissue and may in turn affect melatonin secretion in a paracrine manner. It has been shown that the pineal gland can be induced to express several proinflammatory cytokines [13,15, 16]. The synthesis of inflammatory cytokines in the pineal gland may be induced by LPS interfering with its corresponding receptor‚ÄĒTLR4. The present study showed that expression of TLR4 mRNA in the pineal gland depends on the photoperiodic condition, which confirms results from our previous study that analysed TLRs mRNA expression patterns in the pineal gland of sheep [6]. However, a reduction of TLR4 gene expression after LPS treatment was found in the pineal gland of ewes from the SN photoperiod. The reduction of TLR4 gene expression during inflammatory conditions may be a mechanism that protects pineal cells against extensive stimulation by endotoxins. It is worth noting that our study is based only on the gene expression of TLR4 data. Therefore to better understand the influence of photoperiod on reactivity of pineal gland on the endotoxin, the further study analyzing the expression of TLR4 protein is needed. However, currently it is considered that the expression of the TLRs in the brain is generally variable due to the lack of high specificity antibodies [17]. It is worth noting that other pools of proinflammatory cytokines affecting the secretory activity of the pineal gland could be preset in the cerebrospinal fluid (CSF). The pineal gland is localised in a third ventricle evagination called the pineal recess, and it maintains direct contact with the CSF. It was well established that during peripheral inflammation, proinflammatory cytokines can cross the blood-brain and blood-CSF barriers and reach the CSF [18]. It should be noted that the ability of proinflammatory cytokines to influence melatonin secretion may not exclusively result from the direct action of those mediators on pinealocytes. Immune factors may regulate the secretory activity of pineal gland functioning by affecting pineal glial cells [5]. It was previously found that the action of interferon-gamma (IFN)-ő≥ and IL-1ő≤ on pinealocytes is generally mediated by microglia, whereas TNFőĪ induces its effects by acting directly on pinealocytes because TNF receptors are expressed directly in these cells [16].
Our study suggests that decreased secretion of melatonin after peripheral LPS treatment may result from the reduced synthesis of 5-HT in the pineal gland. Moreover, reduction in the concentration of 5-HT was determined only during the SN photoperiod. This fully reflects the changes in the secretion of melatonin because three hours after treatment, melatonin was lower only in the animals from the SN photoperiod. Although we did not measure the concentration of tryptophan in the pineal gland, it could be concluded that the lowering of pineal 5-HT concentrations in the ewes during the SN photoperiod did not result from a decreased concentration of this amino acid in the tissue, as it was previously found that both LPS treatment [19] and proinflammatory cytokines [20] induced increase of tryptophan concentration in the brain. Our results suggest that lowering 5-HT concentrations in the pineal gland may rather result from a decreased expression of the TPH enzyme, which converts tryptophan to 5-HTP‚ÄĒan intermediate in the pathway of serotonin synthesis. This may significantly influence all downstream stages of melatonin biosynthesis, because TPH1 is characterized by the highest relative level of gene expression among the enzymes of the melatonin biosynthetic pathway. It is worth mentioning that, in contrast to mRNA expression, LPS-dependent suppression of TPH protein expression was determined during both the SN and LN photoperiods. On the other hand, these results suggest that LPS-induced changes in the 5-HT content are not influenced by the other enzyme in the 5-HT synthesis pathway‚ÄĒDDC‚ÄĒbecause no effect of endotoxin treatment was found on the expression of this enzyme. However, an influence of photoperiod on the pineal expression of DDC was found. The reduction of the pineal 5-HT concentration may also result from increased metabolism of this neurotransmitter. Serotonin is metabolised in the pineal gland via two distinct pathways. The first leads to the formation of N-acetylserotonin and then to melatonin. In second one, 5-HT is metabolised by the enzyme monoamine oxidase to yield 5-hydroxyindole acetaldehyde. In turn, this intermediate is then either oxidised to 5-HIAA or reduced to 5-hydroxytryptophol. Both of these are substrates for HIOMT and yield 5-methoxyindole acetic acid and 5-methoxytryptophol, respectively [21]. Our results show that endotoxin-induced inflammation increased the concentration of the 5-HT metabolite‚ÄĒ5-HIAA‚ÄĒin the pineal gland of ewes from both the SN and LN photoperiods. This fully supports the results of the other studies, which also reported increased synthesis of 5-HIAA in the brain during peripheral inflammation and after treatment with IL-1ő≤ [19]. The significant increase in pineal 5-HT metabolism during the LN photoperiod did not influence the total concentration of 5-HT, which suggests that during the LN photoperiod there are the mechanisms regulating pineal 5-HT synthesis that compensate for enhanced degradation of this monoamine. On the other hand, observed independence between the content of 5-HT and 5-HIAA in the pineal gland during the LN photoperiod may be influenced by a different half-life of these two compounds in the brain. The half-life of 5-HIAA is longer than 5-HT, which is degraded in the brain during the first few minutes after formation [22]. Therefore, changes in 5-HIAA may be affected by some amount of metabolite formed during enhanced degradation of 5-HT in the first hour after LPS-treatment, but, due to the fast turnover of 5-HT, this effect was not observed in this monoamine.
Our results also indicate that inflammation may suppress the expression of AANAT in the ovine pineal gland both at the transcriptional and posttranscriptional levels. However, this effect was observed only in the SN photoperiod. In vertebrates, AANAT is considered to be the key enzyme in melatonin synthesis and is known as the melatonin rhythm enzyme. It is currently postulated that the mechanisms regulating melatonin synthesis converge at the control of AANAT enzyme activity [2]. In mammals, NE activates two subtypes of adrenergic receptors located in the pinealocytes leading to the stimulation of melatonin synthesis. Activation of ő≤1-adrenergic receptors increases intracellular concentrations of cAMP, which is followed by the activation of cAMP-dependent protein kinase A (PKA). Both elevated cAMP levels and PKA activation are necessary for the stimulation of AANAT and melatonin synthesis. On the other hand in all mammalian species, activation of őĪ1-adrenergic receptors elevates intracellular calcium ions ([Ca2+]i) levels due to the release of (Ca2+)i from intracellular stores [23]. NE-dependent activation of the ő≤1-adrenergic/cAMP/PKA and őĪ1-adrenergic/(Ca2+)i pathways is conserved in mammalian physiology, but the downstream mechanisms that link these signalling cascades with AANAT activation and melatonin production exhibit marked interspecies variations [2]. In rodents, the cAMP/PKA pathway controls the transcriptional mechanisms regulating melatonin synthesis. However, in ungulates and primates, melatonin synthesis is controlled by mechanisms targeting the posttranslational regulation of AANAT. In these animals, pinealocytes constantly synthesise the AANAT protein from continually available AANAT mRNA. In the absence of noradrenergic stimulation, the AANAT protein is destroyed by proteasomal proteolysis. Under NE stimulation, elevated cAMP levels result in the phosphorylation of AANAT by PKA. This posttranslational modification leads to the interaction of phosphorylated AANAT with 14-3-3 proteins, protecting AANAT from degradation [2]. In our previous ex vivo study on ovine pineal gland explants, however, we determined that treatment with NE increased gene expression of AANAT by approximately 50% compared to the control group [12]. The modulatory role of NE in controlling AANAT transcription in sheep indirectly indicates the occurrence of day/night fluctuations in AANAT mRNA levels in the pineal gland. It was found that the nocturnal expression of AANAT mRNA was approximately 2 times higher than the daytime levels of this transcript [24]. In the present study, it was found that nocturnal expression of AANAT gene is slightly higher during the LN photoperiod compared to the SN photoperiod. However, the changes in gene expression of AANAT were not parallel to the protein expression of this enzyme, which may confirm the importance of posttranscriptional regulation of AANAT expression in sheep. Photoperiod-related differences in the expression of AANAT mRNA may simply be due to higher concentrations of cAMP in the ovine pineal gland during the LN photoperiod compared to the SN photoperiod. However, no influence of inflammation on pineal cAMP concentrations was found regardless of the photoperiod. This suggests that inflammation-induced reductions of AANAT mRNA levels during the SN photoperiod may not result directly from changes in this enzyme‚Äôs gene expression but rather may be an effect of faster posttranscriptional degradation of AANAT mRNA. It was previously found that, at least in rodents, degradation of mRNA encoding AANAT in the pinealocytes is an important mechanism regulating circadian oscillation of this enzyme [25]. However, identifying the factors responsible for enhancing the rate of AANAT mRNA degradation during an immune/inflammatory challenge requires further detailed study. It is worth mentioning that although the activity of AANAT was not examined in our study, an experiment performed on chickens indicated that inflammatory processes besides influencing the expression of AANAT may also reduce activity of this enzyme [4]. It has also been suggested that inflammation may influence melatonin secretion by affecting HIOMT expression and activity. The same study on chickens showed that inflammatory challenges increase HIOMT mRNA expression as well as its enzymatic activity [4]. Additionally, in our ex vivo study on ovine pineal explants, glands collected from LPS-treated animals were characterised by a higher HIOMT expression than those isolated from control ewes [12]. However, in the present study, the inflammatory condition did not influence HIOMT expression regardless of the photoperiod.
It is known that LPS treatment induces not only fever, but also causes transient sickness behaviors (e.g. lethargy, labored breathing and diarrhea) in the ewe [26]. Our study showed that the administration of endotoxin induced stress response in animals regardless of the photoperiodic conditions because the elevated cortisol concentration was determined in blood of all LPS-treated ewes. The stimulatory effect of endotoxin has been previously identified as a strong activator of the hypothalamic-pituitary-adrenal (HPA) stress axis in several different species, including sheep [26,27], and the secretion of melatonin may be influenced by stress. However, the impact of stress on melatonin secretion depends on activity and the diurnal cycle. Stress was found to decrease nocturnal concentrations of melatonin in the pineal gland without affecting AANAT activity in nocturnal animals, such as rats [28], but was found to stimulate nocturnal secretion of melatonin in diurnal ungulate species [29]. However, a study on humans demonstrated that the corticotropin-releasing hormone, a component of the HPA axis, exhibits an inhibitory effect on the pineal secretion of melatonin [30]. In the present study conducted on ewes, the immune stress induced by endotoxin injection suppressed the nocturnal secretion of melatonin, which suggests that during an immune/inflammatory challenge, the role of stress mediators in the modulation of pineal system activity may be abrogated by other inflammatory-derived mediators. It is worth noting that, from the second hour on after treatment, circulating levels of cortisol were higher in the ewes during the LN photoperiod compared to the SN photoperiod. Our results also support results of a previous in vivo study, which showed that melatonin increases the cortisol yield in sheep adrenal glands [31]. Our findings that inflammation more persistently suppressed melatonin secretion in ewes from the SN photoperiod, which was characterised by a lower cortisol concentration after endotoxin treatment, while the inhibitory impact of LPS on melatonin release disappeared one hour after treatment in ewes from the LN photoperiod, which was characterised by higher cortisol concentrations. It suggests that cortisol (and probably other HPA axis mediators) may buffer immune-pineal interactions. It is well established that HPA axis components attenuate the immune response via the inhibition of proinflammatory cytokine synthesis [32]; therefore, increased activity of the HPA axis during the LN photoperiod may at least partially explain faster recovery and normalisation of melatonin secretion in ewes during the LN photoperiod.
CONCLUSION
In summary, our study showed that endotoxin-induced inflammation may disturb the nocturnal secretion of melatonin in ewes, but this effect may be influenced by the photoperiodic conditions. Of great importance is the fact that our research was performed on a non-rodent species‚ÄĒewes. There are significant differences in the intracellular mechanisms regulating melatonin secretion that distinguish rodents and other mammals, including ungulates and primates. Therefore, these results may be considered more universal for day-active species, including humans, than the results of studies conducted on nocturnal animals.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







