





INTRODUCTION
Classical brown adipose tissue (BAT) depots present in the many mammalian fetuses and newborns including humans, sheep, and rodents; and located predominantly around the internal organs and in interscapular and supraclavicular regions [1,2]. In BAT, brown adipocytes contain many small lipid droplets and mitochondria that can uniquely produce large amounts of heat through the activation of non-shivering thermogenesis [3]. This type of thermogenic process is called uncoupled respiration and facilitated by the mediation of its key marker protein, the BAT-specific uncoupling protein uncoupling protein 1 (UCP1) [4,5], which is particularly important for neonatal animals and infants who have greater demands on thermogenesis to adapt to the new extrauterine environment [4,6,7]. As the key window period for sheep fetal BAT development, the BAT growth primarily occurs in the perirenal region over the final third trimester of gestation [8,9]. The BAT cells life cycle starts with differentiation from the preadipocytes expressing Myf5 of embryonic mesoderm and includes a growth phase followed by growth arrest, clonal expansion, and terminal differentiation leading to mature BAT cells [10,11]. In contrast, white adipose tissues (WAT) are derived from the adipogenic lineage, and a white adipocyte contains a single, large lipid droplet for the storage of chemical energy as triglycerides [12]. The predominant type of fat is WAT in adult [13,14].
Fetal undernutrition frequently occurs worldwide including animal agriculture [15]. The intrauterine fetal growth restriction of sheep caused by maternal undernutrition can cause serious decline in the survival rate and growth performance of lambs [16]. In addition, the mortality of newborn lambs caused by hypothermia has accounted for 25% to 35% of the total mortality rate of lambing period, and the number could reach more than 50% in some cases [17]. The appropriate prenatal BAT adipose tissue development is essential for adequate body temperature regulation at birth to ensure immediate survival [8]. The stunted development of BAT has been found in fetal sheep [18], which might be one of the important reasons for the increased mortality of neonatal animals due to their inability to maintain body temperature and resist cold. However, the mechanism underlying the inhibition of fetal BAT growth and development by maternal undernutrition during late pregnancy remains unknown. Therefore, the objective of this study was to investigate the effects of maternal undernutrition during late pregnancy on the regulatory factors of growth and development in ovine fetal perirenal BAT.
MATERIALS AND METHODS
Animals and treatments
All experimental procedures were conducted in conformity with institutional guidelines for the care and use of laboratory animals in China (The State Science and Technology Commission of China, 1988). This study was a companion study of a previous study whose details of animals, experimental design and procedures have been described [19]. In brief, eighteen second or third parity ewes were mated at synchronized estrus and had similar live weights (mean live weights 52.82±2.67 kg) during the d 90 of pregnancy. Pregnancies and singletons were confirmed by ultrasound scanning at approximately d 50 of gestation (Medison-SA-600, Shanghai, China). At the d 90 of pregnancy, the Mongolian ewes were random allocated into three groups (Table 1): restricted group 2 (RG2, 0.18 MJ metabolisable energy [ME]/kg body weight [BW]0.75/d, n = 6), restricted group 1 (RG1, 0.33 MJ ME/kg BW0.75/d, n = 6), and control group (CG, ad libitum, 0.67 MJ ME/kg BW0.75/d, n = 6). All animals were housed in separate pens and fed with chopped hay (mainly Leymus chinensis, Table 2). Change for the experiment was conducted between 90 and 140 days of gestation, as the fetus gained 80% to 85% of its final birth weight during the last two months of pregnancy [20]. Animals were free to drink water and eat mineral mixtures (containing per kilogram: Ca, 15 g; P, 11.5 g; Mg as MgSO4·4H2O, 1 g; Fe as FeSO4·7H2O, 500 mg; Cu as CuSO4·5H2O, 250 mg; Zn as ZnSO4, 175 mg; Mn as MnSO4, 100 mg; Co as CoC12·6H2O, 20 mg; I as KI, 40 mg; Se as Na2SeO3·5H2O, 1.5 mg; Yuan tong weiye Co., Ltd., Inner Mongolian, China). Ewes in the control group were fed at 08:30, 11:00, and 16:00 h per day (The Unconsumed feed accounted for approximately 10% of the total amount provided). The ewes in restricted groups were fed daily at 08:30 and 16:00 h. The unconsumed forage in control group was collected daily and recorded before feeding at 08:30 and sub-sampled for the ME and chemical analysis. The daily intake of hay offered for each group can be calculated by the ewe BW, nutrition value of hay and the energy plane. The formation is below:
DI, daily intake; ME, measured metabolizable energy in hay; L, designed ME level in restricted groups.
Tissue sample collection
At 140 days of pregnancy, the fetuses were collected in from mothers. The body weight and the perirenal fat were measured in fetuses. Some samples of the perirenal fat tissues were snap-frozen in liquid nitrogen and kept at −80°C. Small pieces of the perirenal adipose tissue were fixed with paraformaldehyde (0.1 mol/L, pH 7.4), embedded in paraffin, and cut into 4 to 6 μm sections. Sections were stained with hematoxylin-eosin (H&E) stain kit (D006, NJJCBIO, Nanjing, China) for microscopic examination.
Immunohistochemistry
To detect the expressions of peroxisome proliferator-activated receptor-γ (PPARγ), CCAAT/enhancer-binding protein-α (C/EBPα), and UCP1 in perirenal fat, immunohistochemical staining was performed on 5 μm thick tissue sections using rabbit polyclonal antibodies: PPARγ (bs4590R, BIOSS, Bengjing, China), C/EBP alpha (GTX100674, Gene Tex, Alton Pkwy, CA, USA), UCP1 (ab10983, Abcam, Waltham, MA, USA) as the first antibody in phosphate buffered saline-Tween. The secondary antibody was a horseradish peroxidase (HRP) (ab205718, Abcam, USA). Thermally induced antigen retrieval was performed by incubation with Tris-ethylene diamine tetraacetic acid for 20 minutes. Endogenous peroxidase was blocked by incubation with 3% H2O2 for 10 minutes. Non-specific binding was blocked by incubating sections in normal goat serum for 2 hours at room temperature. Visualization was performed by incubation with HRP conjugated streptavidin followed by staining with diaminobenzidine substrate. Images were analyzed using a Leica microscope (Leica, DM750, Wetzlar, Germany) equipped with a ICC50 W camera (Leica) and Leica Application Suite software (version 4.6.2) at a magnification of 400×. Image J (v1.45) software was used to determine the area of the positive signal divided by the total tissue area to calculate the area fraction of the positive signal.
Immunoblotting and antibodies
Total protein was extracted by lysing cells with RIPA lysate I (C5000050010; Sangon Biotech, Shanghai, China). Protein concentration was measured using a BCA Protein Assay Kit (C5030210500; Sangon Biotech, China). Equal amounts of protein lysate were boiled for 5 minutes with 5× sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) loading buffer (P1040; Solarbio, Beijing, China) and separated by SDS-PAGE Preparation kit (C631100; Sangon Biotech, China) and transferred to polyvinylidene fluoride membrane (Millipore ImmobilonTM P, Billerica, MA, USA). After blocking in 5% skim milk dissolved in Trisbuffered saline (0.02 M Tris base, 0.14 M NaCl, pH 7.4) containing 0.1% Tween 20, the membrane was incubated with primary antibodies overnight at 4°C, and then with IRDye 800CW goat anti-rabbit immunoglobulin G (Li-COR Biotechnology, Lincoln, NE, USA) for 1 hour at room temperature. The primary antibodies used in this study were UCP1 (ab10983; Abcam, USA), myf5 (ab139523; Abcam, USA), PPARγ (bs4590R; Bioss, China), CEBPα (GTX100674; Gene Tex, USA), β-actin (bs0061R; Bioss, China) and glyceraldehyde-3-phosphate dehydrogenase (HC301; TransGen, Beijing, China). The blots were imaged by the Odyssey CLx Imaging System (Li-COR Biotechnology, USA) and the protein density analysis was performed using Image J (v1.45) software.
Real-time polymerase chain reaction analysis
Total RNA was isolated using RNAiso Plus reagent (9109; Takara, Shiga, Japan) according to the manufacturer’s protocol. Reverse transcription of total RNA (500 ng) was performed using a PrimeScript RT reagent Kit (RR047B; Takara, Japan). Real-time reverse transcriptase (RT)-polymerase chain reaction (PCR) was performed using SYBR Premix Ex Taq (RR820A; Takara, Japan) in a LightCycler480 PCR system (Roche, Basel, Switzerland). The thermal cycling parameters were as follows: 95°C for 30 s for initial denaturation then 40 cycles of 95°C for 5 s, 60°C for 30 s and 72°C for 30 s. Reactions were run in triplicate for each sample. Primer sequences used in RT-PCR of selected genes are provided in Table 3. The cDNA of perirenal adipose tissue was diluted with 10 times concentration gradient for RT-PCR according to the above conditions and systems, and then the logarithm of template initial concentration was plotted with Ct to obtain the standard curve of each gene. At the same time, the amplification curve and dissolution curve of each gene in the process were recorded to judge the unity and integrity of the amplification product. The standard curve is controlled by amplification efficiency (Amplification efficiency = 10−1/slope) and correlation coefficient. All samples were measured in triplicate, and the real-time PCR assay had similar efficiency and within the range of 90% to 110%. Threshold cycle (Ct) was the value of PCR cycles at which the fluorescence signal of the PCR reaction reached a fixed threshold. For each sample, the Ct both for the target gene and endogenous control gene were determined to calculate ΔCt sample (Cttarget gene – Ctendogenous control). In this study, the β-actin gene was used as an endogenous control. Subsequently, ΔΔCt (ΔCtsample – ΔCtCG) was determined, and the relative expression was calculated by 2−ΔΔCt. For the CG group, the ΔΔCt is equal to 0. Negative controls were performed in which cDNA was substituted by water.
RESULTS
Fetal perirenal fat growth and structure
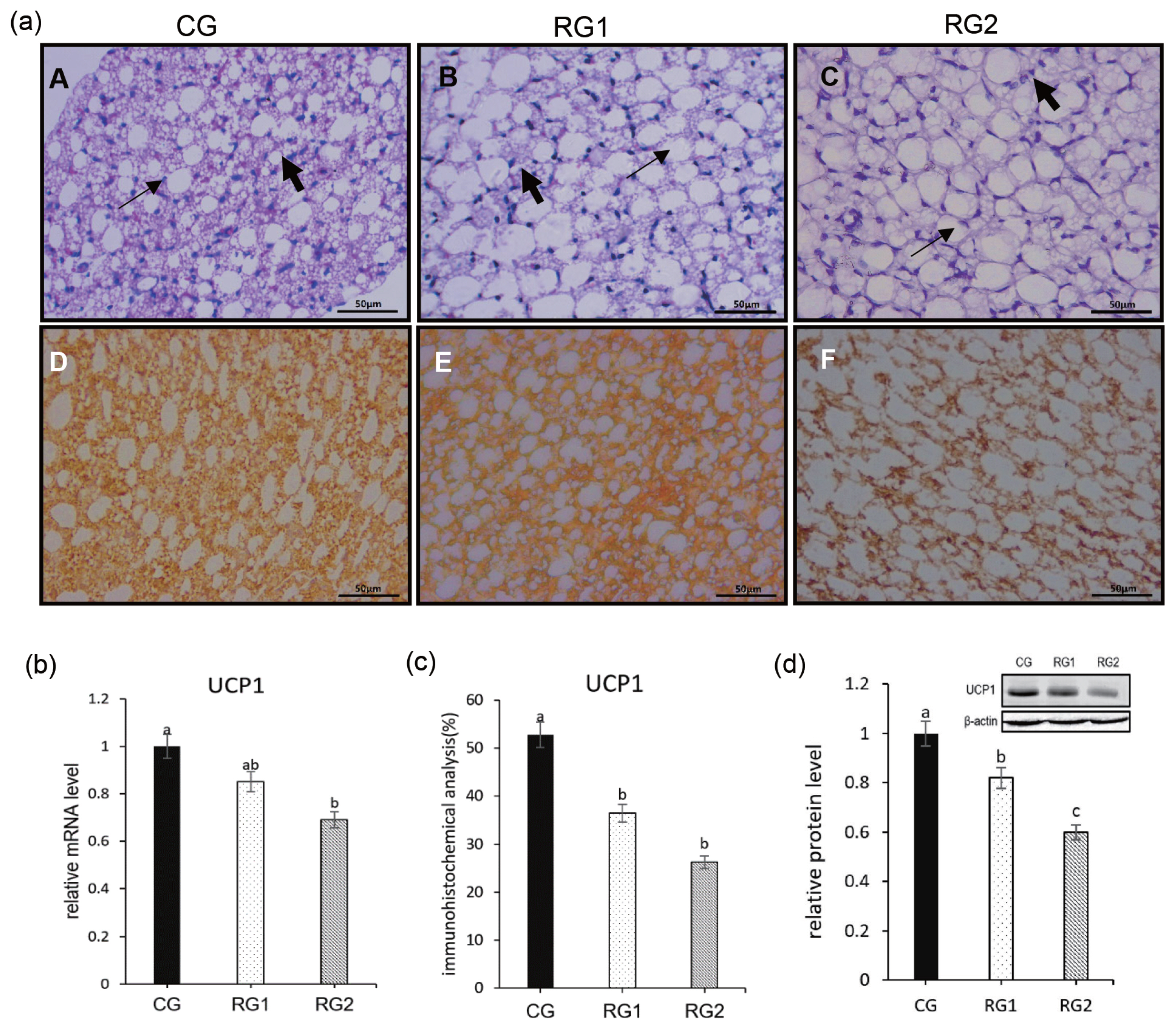
With the reduction of maternal feed intake, the fetal body weight was reduced in RG2 and RG1 (p = 0.0069), and fetal perirenal fat weight (p = 0.0077) and perirenal fat growth rate (p = 0.0074) were decreased in RG2 group compared to CG group. For the ratio of perirenal fat to body weight, there were no differences between the restricted groups and the CG group (p = 0.0613) (Table 4). In the restricted groups, the number of WAT cells increased gradually with the decrease of the energy intake and the lipid droplets became larger, whereas BAT cells became sparse compared to control group from H&E staining of sections of fetal perinatal adipose tissues. As the nutrient intake decreased, UCP1 protein level was significantly lower in the RG2 and RG1 compared with the control group (p = 0.0001). The relative mRNA expression of UCP1 was decreased in the RG2 compared with CG and RG1 (p = 0.0265). In addition, a decrease (p = 0.0008) in the immunostaining of UCP1 was seen in RG2 and RG1 groups (Figure 1).
Myf5 expression in fetal perirenal brown adipose tissue
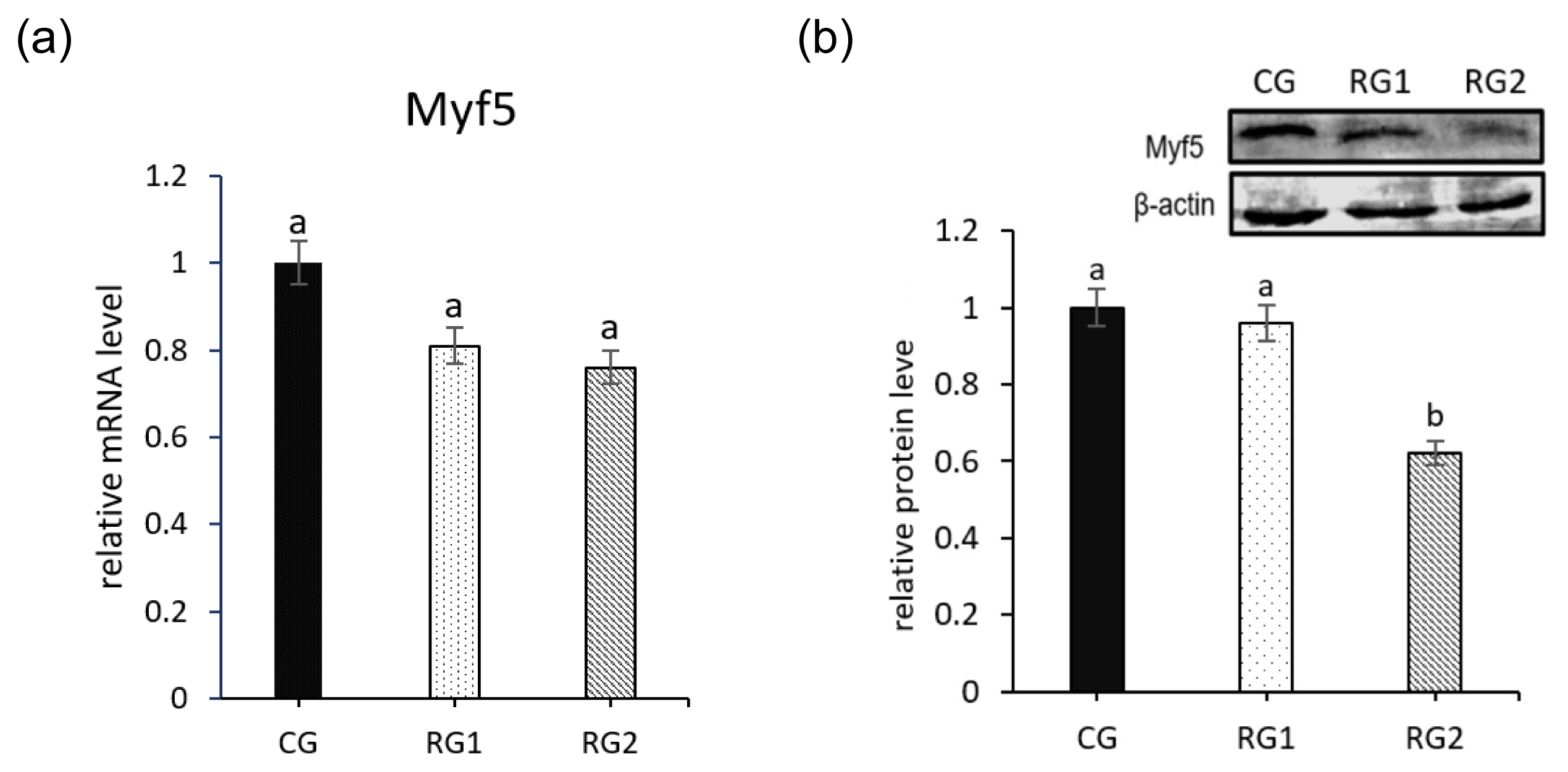
Effects of feed restriction during late pregnancy on the gene expression of Myf5 in fetal perirenal BAT are shown in Figure 2. No significant difference was seen in mRNA expression of Myf5 in restricted groups (p = 0.3100). However, the relative protein level of Myf5 were decreased in RG2 group compared with the CG group (p = 0.0001).
Expression of cell cycle regulatory factors in fetal perirenal brown adipose tissue
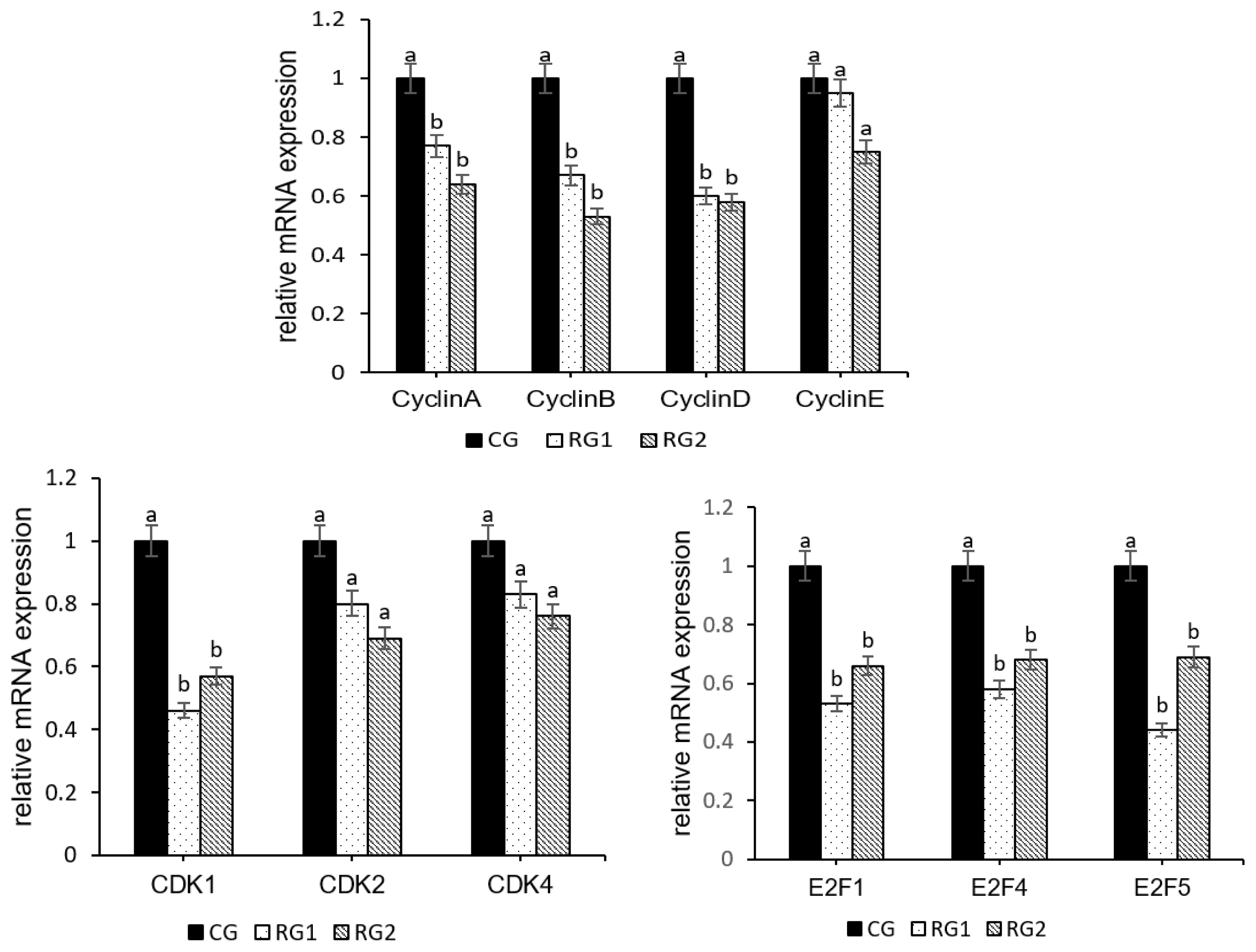
As show in Figure 3, the mRNA expressions of CyclinA (p = 0.0109), CyclinB (p = 0.0019), CyclinD (p = 0.0015), CDK1 (p = 0.0001), E2F1 (p = 0.0323), E2F4 (p = 0.0101), and E2F5 (p = 0.0018) were lower in the RG2 and RG1 groups than those in the CG group, however, no differences were observed in expressions of cyclin-dependent kinase 2 (CDK2) (p = 0.1608), CyclinE (p = 0.2847), and cyclin-dependent kinase 4 (CDK4) (p = 0.4074) among the fetal peripheral BAT of three groups.
Expression of PPARγ and C/EBPα in fetal perirenal brown adipose tissue
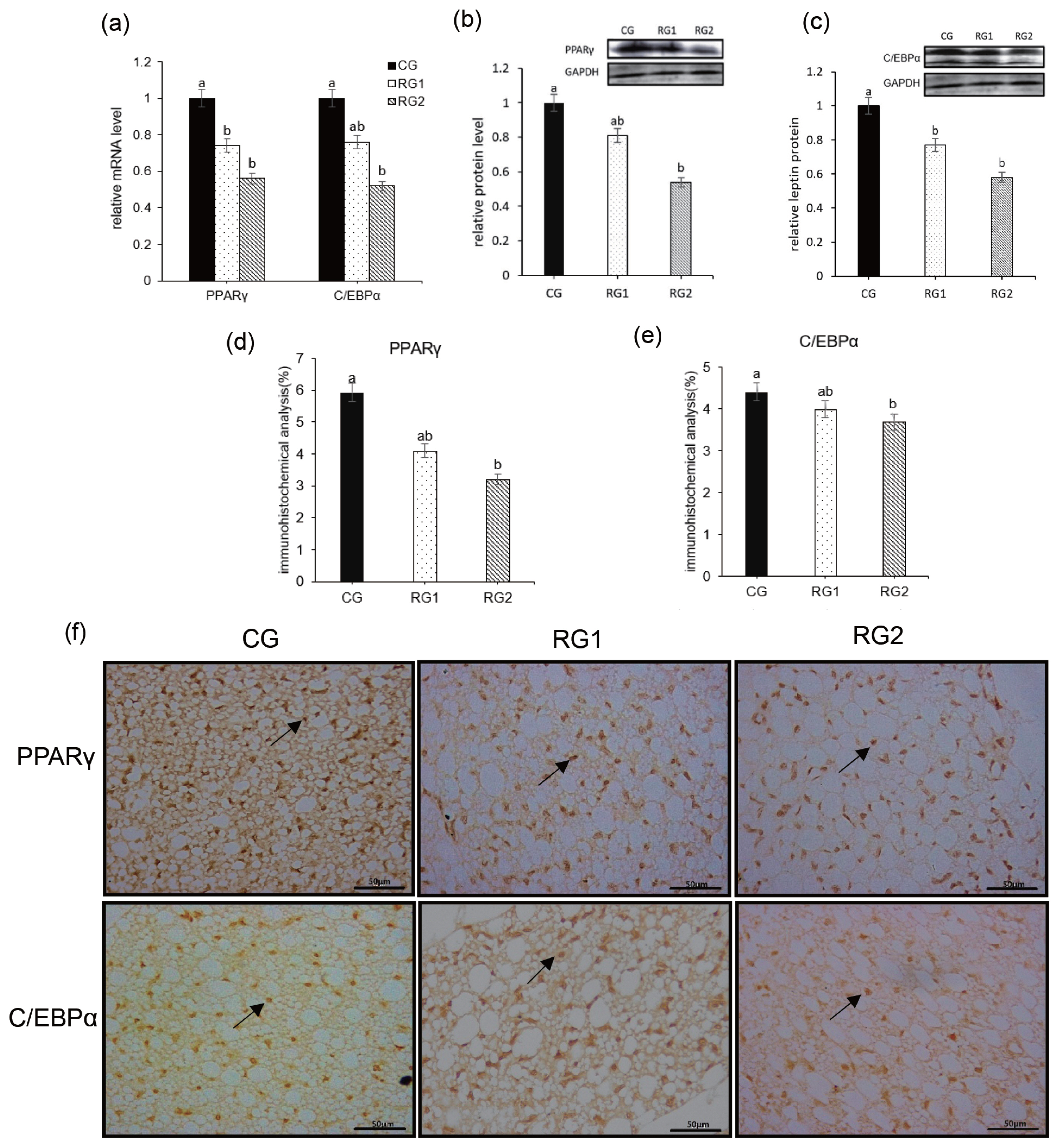
Figure 4 shows the effect of feed restriction during late pregnancy on the expression of PPARγ and C/EBPα in fetal peripheral adipose tissue. As the nutrient intake decreased, the relative mRNA expression of C/EBPα in the RG2 group was significantly lower than that of the CG group (p = 0.0307) and the relative protein levels were reduced in both restriction groups (p = 0.0015). Consistent with this, the reduced mRNA expression of PPARγ (p = 0.0008) was found in RG2 and RG1 groups, and the relative protein expression of PPARγ (p = 0.0043) was decreased in RG2 group compared to the CG group. Moreover, the decreased immunostaining of PPARγ (p = 0.0341) and C/EBPα (p = 0.0076) was seen in the RG2 and RG1 groups.
Expression of others regulatory factors in fetal perirenal brown adipose tissue
Effect of feed restriction during late pregnancy on the gene expression of others regulatory factors in fetal peripheral BAT is presented in Table 5. Expression of BMP4 (p = 0.0083) and BMP7 (p = 0.0330) mRNA decreased in RG2 group compared to CG group, but there was no difference found in the EBF2 gene expression (p = 0.6757). The mRNA expression of peroxisome proliferator-activated receptor co-activator-1α (PGC-1α) reduced in both RG2 and RG1 groups (p = 0.0104). Furthermore, there was no significant difference in the gene expression of positive regulatory domain-containing protein 16 (PRDM16) (p = 0.6146), krüppel-like factor 4 (KLF4) (p = 0.4275), GATA binding protein 2 (GATA2) (p = 0.0897), and transforming growth factor type-β (TGFβ) (p = 0.1223) among those groups.
DISCUSSION
Adipose tissue is obviously limited by nutrition, and maternal changes can significantly change the development of fetal fat [21]. In fetus, the adipocyte progenitor cell line of BAT is gradually transformed from mesenchymal stem cells (MSCs) through a complex series of differentiations [22]. Adipogenesis is the process of cell differentiation from preadipocytes to adipocytes, and the initial phase of BAT adipogenesis is characterized by the proliferation of Myf5 preadipocytes [11]. In addition, preadipocytes may be retained with the potential to develop into brown adipocytes even after brown adipocytes disappear [23]. During those processes, the cells expressing Myf5 are the direct preadipocytes of BAT although Myf5 is a myogenic regulator [24]. In this study, the Myf5 protein expression was significantly decreased in the RG2 group compared to the CG group, suggesting that the formation of the BAT adipocyte progenitor cell line in the RG2 group may be repressed by maternal under nutrition. Furthermore, the mRNA and protein expression of ovine fetal perirenal UCP1, a biomarker of BAT, was decreased in restricted groups, consistent with the reduced Myf5 protein in RG2 fetal perirenal BAT. The reduced Myf5 preadipocytes of BAT may be one of the important reasons that cause decreased maturation of fetal perirenal BAT and weight of perirenal BAT in the restricted groups.
Preadipocytes enter the cell cycle and experience a growth phase, until they eventually exit the cell cycle and acquire the features of mature BAT adipocytes [25]. The mature adipocytes don’t have the ability to proliferate but with the ability for expansion, thus, the number of mature BAT adipocytes depends on the proliferation regulated by cell cycling of the Myf5 preadipocytes [11]. The proliferation of preadipocytes regulates the completion of cell cycle through the joint action of cyclins and cyclin dependent kinases, which are vital regulators for the normal cell cycling [26]. CDK4 and CyclinD are responsible for getting cells out of quiescence (G0) and driving them through G1 [27]. The reduced gene expression of CyclinD and tended decrease in CDK4 seen in our study may indicate that the G1 phase development of BAT precursor cells was blocked. CDK2-CyclinA and CyclinE-CDK2 complexes are essential for completing S phase correctly [28], whereas CDK1-CyclinB is a vital regulator for normal withdrawal from mitosis [29]. In addition, E2Fs play roles that contribute to timely progression through G1/S phases [30]. The reduced gene expression of CDK1, CyclinB, E2F1, E2F4, and E2F5 in the restriction group in this study suggests that DNA synthesis and mitosis were inhibited during the proliferation of fetal perirenal BAT preadipocytes. Collectively, the inhibition of proliferation of preadipocytes in the present study may be directly attributed to variation of cell cycle regulators.
The BAT preadipocytes differentiate into mature adipocytes [31]. Adipogenic differentiation transcriptional cascades that mainly involve PPAR-γ and the C/EBPα are considered the crucial determinants of adipocyte fate [32,33]. In our study, PPARγ mRNA expression and C/EBPα protein expression in perirenal fat were lower in the RG2 and RG1 groups compared with the control group, the PPARγ protein expression and C/EBPα mRNA expression were decreased in RG2 fetuses. These decreases might also reduce preadipocyte development to mature BAT adipocytes and contribute to the decrease of perirenal fat mass. PPARγ gene expression was found to positively correlate to perirenal fat mass and birth weight [34]. While PPARγ and C/EBPα are well known essential transcriptional modulators of adipocyte development in all types of adipose tissue [35], EBF2, BMP7, BMP4, PRDM16, PGC-1α, KLF4, GATA2, and TGFβ also regulate adipogenesis [36]. As a pioneer factor in guiding and promoting the fate of BAT precursor cells in the process of MSCs differentiation into BAT potential precursor cells [37,38], the expression of EBF2 decreases with reduction of maternal nutrition, inhibits the development of BAT precursor cells, and reduces UCP1 expression [38]. BMP7 and BMP4 promote the acquisition of brown phenotype and regulate EBF2 activity [39]. BMP7 and BMP4 gene expressions were decreased in the RG2 group in this study, which may contribute to inhibition of the acquisition of BAT adipocyte phenotype in maternal malnutrition and hinder the normal differentiation of BAT preadipocytes in fetal perirenal fat. Meanwhile, PPARγ and C/EBPα initiate positive feedback to induce their own expression and activate many target genes whose expression determines the BAT adipocyte [40]. PRDM16 stimulates the key characteristics of authentic brown adipocytes [41]. In addition, PGC-1α plays a central role in BAT gene expression and coordinates the gene expression of key components of mitochondrial biogenesis in BAT [42]. KLF4 is required for promoted WAT differentiation, and its increase inhibits the development of BAT [14]. GATA2 expression blunts adipocyte differentiation through reduced PPARγ activity [43], whereas TGF-β inhibits the production of adipose precursor cells which reduces the opportunity for the formation of mature BAT cells [44]. In this study, the gene expression of PGC-1α decreased in the restricted groups and PRDM16 showed a downward trend, while the KLF4 was increased, and no significant differences were found in GATA2 and TGF-β. These results indicate that inhibition of PPARγ and C/EBPα expression is associated with a decrease in the expression of other transcription target genes involved in adipogenesis in fetal perinephric BAT depot.
CONCLUSION
The reduction of BAT precursor cell cycle regulators causes the retardation of preadipocytes and followed by the decreased expression of BAT precursor cell development regulators including BMP4 and BMP7. Ultimately the expressions of PPARγ, C/EBPα, and PGC-1α promoting differentiation were decreased which resulted in the inhibition of BAT maturation in fetal perirenal fat during late pregnancy with maternal undernutrition. These changes would be one of crucial potential-mechanisms hypothermia and high mortality of new born lambs.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print







